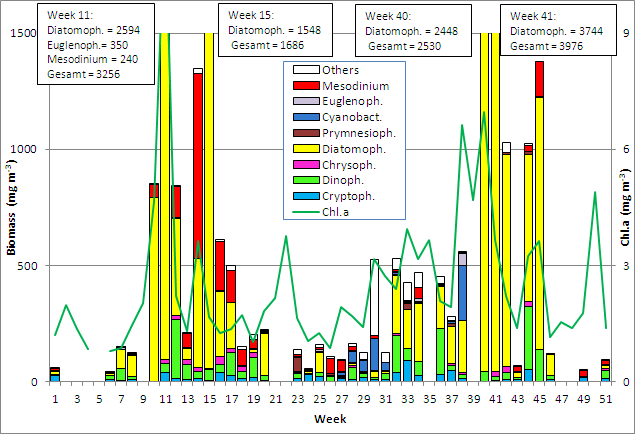
Phytoplankton development at the coastal station "Seebrücke Heiligendamm" in 2010
The Leibniz Institute for Baltic Sea Research conducts a coastal monitoring programme with weekly samplings at the sea-bridge Heiligendamm (54°08,55' N; 11°50,60' E; 300 m off shore, 3 m water depth). The Department of Marine Biology analyses the surface samples, taken by means of a bucket, for phytoplankton composition and biomass and for chlorophyll a.
The phytoplankton biomass is determined by microscopical counting (UTERMÖHL method) and the chlorophyll a concentration by ethanol extraction and fluorometric measurement. Method instructions see http://www.helcom.fi/groups/monas/CombineManual/AnnexesC/en_GB/.
Phytoplankton counting was carried through by use of the counting programme OrgaCount and is based on the HELCOM-biovolume factors which are annually updated: http://www.ices.dk/env/repfor/index.asp (basics see in Olenina et al. 2006). The analytical specifics of the chlorophyll a determination are published by Wasmund et al. (2006). According to the decision of the BLMP-subgroup “Quality Assurance” from 11.9.2008 we show here chlorophyll a data which are not corrected for pheopigments.
Microscopical analysis was not possible in 12 samples because of high sediment portions, caused by wind-induced sediment resuspension at the shallow station. Chlorophyll measurements were possible, except for week 6.
As expected, the phytoplankton biomass was very low in the first few weeks of the year. The chlorophyll a data suggest that during the period of sample shortfall until 9.2.2010 no significant phytoplankton growth occurred. From the 16.2.2010 (Week 7) onwards, diatoms developed as already found in 2009, however not by Thalassiosira spp. ![]() but by Rhizosolenia setigera (Image 1) and as usual by Skeletonema costatum .
but by Rhizosolenia setigera (Image 1) and as usual by Skeletonema costatum .

In contrast to the year 2009, not only Skeletonema costatum but several diatom species constituted the main components of the bloom, which occurred since week 10. The spring bloom consisted at its peak (16.3.2010, week 11) mainly of Rhizosolenia setigera (801 mg m-3), Skeletonema costatum (787 mg m-3), Chaetoceros wighamii (617 mg m-3) ![]() and Thalassiosira spp. (253 mg m-3).
and Thalassiosira spp. (253 mg m-3).
Surprising was the spontaneous appearance of the euglenophyte Eutreptiella sp. (350 mg m-3) ![]() , which was also found in 2008. At the same time, chlorophyll a concentrations reached the annual maximum (12.25 mg m-3 ). That means: We found in 2010, just like in 2009, a very early spring bloom which contained 80% diatom biomass, and it was rather diverse.
, which was also found in 2008. At the same time, chlorophyll a concentrations reached the annual maximum (12.25 mg m-3 ). That means: We found in 2010, just like in 2009, a very early spring bloom which contained 80% diatom biomass, and it was rather diverse.
The bloom declined quickly by week 13, while primarily heterotrophic dinoflagellates (Protoperidinium pellucidum et spp. ; Gymnodiniales) developed.
The diatoms reappeared in week 14./15., accompanied by high biomass of the ciliate Mesodinium rubrum . This second diatom development was at first dominated by Skeletonema costatum (333 mg m-3 in week 15) as expected. Astonishingly, Skeletonema costatum disappeared almost completely by week 15 (13.4.2010) and was replaced by a bloom of Coscinodiscus radiatus (966 mg m-3, Image 2). The diatom bloom broke down completely by the 4.5.2010, but also Mesodinium rubrum was reduced whereas heterotrophic flagellates developed, obviously on the basis of the organic remains of the bloom. On 18.5.2010, Coscinodiscus radiatus (Image 2) dominated again.
It is exciting that Dictyochophyceen were only weakly developed. They are shown in Fig. 1 under the group of Chrysophyceae, which did not exceed 35 mg m-3 in the year 2010. The silicoflagellate Dictyocha speculum appeared in the years 2007 and 2008 even abnormally early. This special situation of the years 2007 and 2008 did not continue in 2009 and 2010.

The quick changes of the dominating species cannot be caused by extreme growth rates but it can be explained by the drifting of different water bodies containing different species composition along our fixed station. It allows insight into the heterogeneity of the sea area and the diversity in community structure.
From 8.6. to 3.8.2010, the phytoplankton was low in
biomass but high in diversity. The short appearance of the potentially toxic Chrysochromulina sp. on 8.6.2010 is
noteworthy (Image 3). The spring species Coscinodiscus
radiatus and Skeletonema costatum
gained again dominance on 22.6.2010 (week 25). The chlorophyll a concentrations exceeded 3 mg m-3
only on 27.7.2010 (week 30) within the period from 8.6. to 3.8.2010, when cyanobacteria
reached their summer maximum (Nodularia
spumigena ![]() : 57 mg m-3; Aphanizomenon sp.
: 57 mg m-3; Aphanizomenon sp. ![]() : 79 mg m-3).
Noteworthy is the occurrence of the Prasinophyte Pyramimonas sp. (Image 4), which is counted to „Others“ in Fig. 1.
: 79 mg m-3).
Noteworthy is the occurrence of the Prasinophyte Pyramimonas sp. (Image 4), which is counted to „Others“ in Fig. 1.


On 10.8.2010 (Week 32), the next population change occurred: the dinoflagellate Alexandrium pseudogonyaulax (140 mg m-3, Image 5) and the typical summer diatom Dactyliosolen fragilissimus ![]() (113 mg m-3). appeared. It was the first identified record of Alexandrium pseudogonyaulax in the IOW’s sample history.
(113 mg m-3). appeared. It was the first identified record of Alexandrium pseudogonyaulax in the IOW’s sample history.

The typical autumn diatom Coscinodiscus granii ![]() became dominant on 17.8.2010, it continued growth besides of Dactyliosolen fragilissimus
became dominant on 17.8.2010, it continued growth besides of Dactyliosolen fragilissimus ![]() and stayed the dominant diatom until 21.9.2010 (week 38). The typical dinoflagellate of autumn, Ceratium spp., as well as the diatom Melosira moniliformis
and stayed the dominant diatom until 21.9.2010 (week 38). The typical dinoflagellate of autumn, Ceratium spp., as well as the diatom Melosira moniliformis ![]() developed in parallel.The occurrence of the summer cyanobacteria Nodularia spumigena
developed in parallel.The occurrence of the summer cyanobacteria Nodularia spumigena ![]() (236 mg m-3) at this late date is extremely unusual.
(236 mg m-3) at this late date is extremely unusual.
Like in autum 2009, diatoms dominated also in autumn 2010, whereas the otherwise bloom-forming dinoflagellates (Ceratium spp.) were almost completely absent. The most important diatoms on 5.10.2010 (week 40) were: Chaetoceros socialis (793 mg m-3, Image 6), Cerataulina pelagica (418 mg m-3) ![]() , Pseudosolenia calcar-avis (376 mg m-3, Image 7), Pseudo-nitzschia spp. (324 mg m-3) and Chaetoceros curvistus (135 mg m-3)
, Pseudosolenia calcar-avis (376 mg m-3, Image 7), Pseudo-nitzschia spp. (324 mg m-3) and Chaetoceros curvistus (135 mg m-3) ![]() . The mass occurrence of Chaetoceros socialis is unusual. Pseudosolenia calcar-avis is new in the IOW’s species list. The peak was reached on 12.10.2010 with Cerataulina pelagica (1844 mg m-3), Pseudosolenia calcar-avis (684 mg m-3) and Pseudo-nitzschia spp. (673 mg m-3). This bloom remained at least until 9.11.2010 (week 45). The minima in weeks 43 und 46 must not be considered for quantitative analyses because microscopical countings were not possible owing to a high share of sediment particles; they were performed only for a qualitative overview. The samples from week 47 onwards were completely not usable for quantitative analyses.
. The mass occurrence of Chaetoceros socialis is unusual. Pseudosolenia calcar-avis is new in the IOW’s species list. The peak was reached on 12.10.2010 with Cerataulina pelagica (1844 mg m-3), Pseudosolenia calcar-avis (684 mg m-3) and Pseudo-nitzschia spp. (673 mg m-3). This bloom remained at least until 9.11.2010 (week 45). The minima in weeks 43 und 46 must not be considered for quantitative analyses because microscopical countings were not possible owing to a high share of sediment particles; they were performed only for a qualitative overview. The samples from week 47 onwards were completely not usable for quantitative analyses.


An unusually late diatom autumn bloom, as found in 2009, could not be confirmed because of non-analyzable samples. The relatively high chlorophyll a concentration on 14.12.2010 (week 50) could be an indication of a relatively late bloom. Already in the year 2008, a bloom of Coscinodiscus granii ![]() appeared as late as 16.12.08.
appeared as late as 16.12.08.
If the phenomena of earlier spring blooms and later autumn blooms will be confirmed in future, this indicates a prolongation of the vegetation period, which might be caused by the global tendency of warming.
Literature
Olenina, I., Hajdu, S., Andersson, A.,Edler, L., Wasmund, N., Busch, S., Göbel, J., Gromisz, S., Huseby, S., Huttunen, M., Jaanus, A., Kokkonen, P., Ledaine, I., Niemkiewicz, E. (2006): Biovolumes and size-classes of phytoplankton in the Baltic Sea. Baltic Sea Environment Proceedings No.106, 144pp.
Internet-Zugang:
Paper: http://www.helcom.fi/stc/files/Publications/Proceedings/bsep106.pdf
Tabelle: http://www.helcom.fi/stc/files/Publications/Proceedings/
bsep106ANNEX1Biovolumes_web.xls
Wasmund, N., Topp, I., Schories, D. (2006): Optimising the storage and extraction of chlorophyll samples. Oceanologia 48: 125-144.
IOW, 10.02.2010
Dr. Norbert Wasmund,
Susanne Busch,
Ina-Marie Topp,
Regina Hansen.

State of the Baltic Sea
- Annual Reports on the state of the Baltic Sea Environment
- Cruise Reports
- Data from the autonomous measuring stations
- Development of the suboxic and anoxic regions since 1969
- Baltic Thalweg transect since 2014
- Algal blooms at Heiligendamm since 1998
- "Major Baltic Inflow" December 2014
- "Major Baltic Inflow" January 2003
- Baltic saline barotropic inflows 1887 - 2018
- Further Reading