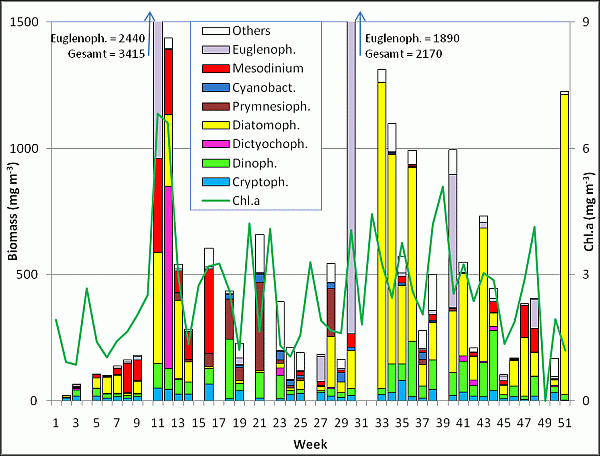
Phytoplankton development at the coastal station "Seebrücke Heiligendamm" in 2008
The Leibniz Institute for Baltic Sea Research conducts a coastal monitoring programme with weekly samplings at the sea-bridge Heiligendamm (54°08,55' N; 11°50,60' E; 300 m off shore, 3 m water depth). The Department of Marine Biology analyses the surface samples, taken by means of a bucket, for phytoplankton composition and biomass and for chlorophyll a.
The phytoplankton biomass is determined by microscopical counting (UTERMÖHL method) and the chlorophyll-a-concentration by ethanol extraction and fluorometric measurement (calculation according to LORENZEN method). Method instructions see
(http://www.helcom.fi/Monas/CombineManual2/CombineHome.htm).
For phytoplankton counting, the program PhytoWin and the HELCOM-biovolume factors (Olenina et al. 2006) were used. The analytical specifics of the chlorophyll-a-determination are published by Wasmund et al. (2006).
Microscopical analysis was not possible in 7 samples because of high sediment portions, caused by wind-induced sediment resuspension at the shallow station. Chlorophyll measurements were, however, always possible.
The phytoplankton biomass was very low in the first few weeks of the year. It was mainly composed of cryptophyceae (Plagioselmis prolonga, Teleaulax spp., Hemiselmis spp.) on 8.1.08. Since week 3 (15.1.08), Gymnodiniales appeared, and since week 5 (29.1.08) also Ceratium tripos and Porosira glacialis. Remarkably, a quick change within the diatoms occurred from Porosira glacialis to Rhizosolenia setigera from week 5 to 6, and back to Porosira glacialis in week 7. Obviously, different water bodies with different species signature drifted along the station. Since week 7 (12.2.08), Mesodinium rubrum developed and reached a biomass of 118 mg m-3 in week 8. On 26.2.08 (week 9), Mesodinium rubrum, Porosira glacialis, Eutreptiella sp., Teleaulax spp. and Guinardia delicatula contributed more than 10 mg m-3 in each case. Unfortunately, no detailed information on the start of the spring bloom was possible because the sample from 4.3.08 could not be evaluated. On 11.3.08, the spring bloom has reached already its peak (3415 mg m-3). Astonishingly, it was not dominated by diatoms but by the euglenophyceae Eutreptiella spp. (2440 mg m-3). Besides this, the diatoms Rhizosolenia setigera and Rhizosolenia hebetata forma semispina, and the photoautotrophic ciliate Mesodinium rubrum were important. Also Teleaulax spp., Gyrodinium spirale, Protoperidinium depressum, Ceratium tripos and Proboscia alata are worth mentioning.
The quick disappearance of the spring bloom might have been affected by a transport of water bodies because the species composition differed very much from the 11.3.08 (wek 11) to the 18.3.08 (week 12). Eutreptiella spp. was suddenly almost completely absent, whereas the silicoflagellate Dictyocha speculum appeared with 722 mg m-3. As it occured mainly in its "naked" form, its identification is not sure; also the genera Chattonella, Verrucophora or Pseudochattonella could be possible. Important diatoms were Rhizosolenia hebetata forma semispina, Proboscia alata, Rhizosolenia setigera and Guinardia delicatula.
By the 25.3.08 (week 13), also Dictyocha speculum disappeared. Suddenly, Skeletonema costatum became dominant (270 mg m-3). It is regularly one of the most important spring diatoms. Despite a taxonomic revision of the Skeletonema-species (vgl. Sarno et al., 2005) we keep the old name Skeletonema costatum preliminarily.
The diatoms vanished completely by the 15.4.08 (week 16). Mixotrophic organisms appeared, like cryptophyceae (Plagioselmis prolonga) and prymnesiophyceae (Chrysochromulina spp.). Perhaps also the dominating ciliate Mesodinium rubrum belongs to this mixotrophic group.
The next change occurred until the 29.4.08 (week 18): Mesodinium rubrum and Cryptophyceae were absent, but Chrysochromulina spp. developed. Dinoflagellates (Heterocapsa triquetra, Heterocapsa rotundata) became dominant. The early appearance of Aphanizomenon sp. (12 mg m-3) is worth mentioning. The phytoplankton composition was similar on 20.5.08 (week 21).
On 3.6.08 (week 23), Chrysochromulina spp. has vanished. Small unidentified unicells dominated. Perhaps also small hardly recognizable Chrysochromulina spp. were counted into this group, but it was mainly composed of coccoid cyanobacteria.
The phytoplankton biomass declined further by the end of June. The dominating "Unidentified" were mainly made up by small coccoid cyanobacteria (< 2 µm) and very thin filamentous cyanobacteria (< 1 µm). The last sample of June could not be counted because of too much sediment in the sample.
Our data from the first half of the year 2008 showed again that this coastal station is not optimal for recording the phytoplankton succession. The general phytoplankton development is well reflected (e.g. timing of the bloom, magnitude of biomass). However, abrupt changes in species composition cannot be considered in the sense of succession. They are based on macro-scale differences in species composition which become apparent if the different water bodies are drifting along the fixed station. If the phytoplankton data of this one station would be combined with data on water currents, they would allow to describe even spatial heterogeneities.
Literature
Olenina, I., Hajdu, S., Andersson, A.,Edler, L., Wasmund, N., Busch, S., Göbel, J., Gromisz, S., Huseby, S., Huttunen, M., Jaanus, A., Kokkonen, P., Ledaine, I., Niemkiewicz, E. (2006): Biovolumes and size-classes of phytoplankton in the Baltic Sea. Baltic Sea Environment Proceedings No.106, 144pp.
Internet-Zugang:
Paper: http://www.helcom.fi/stc/files/Publications/Proceedings/bsep106.pdf
Tabelle: http://www.helcom.fi/stc/files/Publications/Proceedings/bsep106ANNEX1Biovolumes_web.xls
Sarno, D., Kooistra, W.H.C.F., Medlin, L., Percopo, I., Zingone, A. (2005). Diversity in the genus Skeletonema (Bacillariophyceae). II. An assessment of the taxonomy of S. costatum-like species with the description of four new species. J. Phycol. 41, 151-176.
Wasmund, N., Topp, I., Schories, D. (2006): Optimising the storage and extraction of chlorophyll samples. Oceanologia 48: 125-144. Wasmund, N., Pollehne, F.; Postel, L., Siegel, H., Zettler, M.L. (2008): Biologische Zustandseinschätzung der Ostsee im Jahre 2007. Meereswiss. Ber., Warnemünde, 74, 88 pp.
IOW, 29.01.2009
Dr. Norbert Wasmund,
Susanne Busch,
Ina-Marie Topp,
Regina Hansen.

State of the Baltic Sea
- Annual Reports on the state of the Baltic Sea Environment
- Cruise Reports
- Data from the autonomous measuring stations
- Development of the suboxic and anoxic regions since 1969
- Baltic Thalweg transect since 2014
- Algal blooms at Heiligendamm since 1998
- "Major Baltic Inflow" December 2014
- "Major Baltic Inflow" January 2003
- Baltic saline barotropic inflows 1887 - 2018
- Further Reading