

Phytoplankton an der Küstenstation "Seebrücke Heiligendamm" im Jahre 2008
Die Sektionen Biologische Meereskunde und Meereschemie des Leibniz-Instituts für Ostseeforschung Warnemünde (IOW) führen wöchentliche Probennahmen an der Seebrücke Heiligendamm (54°08,55' N; 11°50,60' E) durch. Hier werden die Ergebnisse der Phytoplankton-Untersuchungen vorgestellt. Es wurde Oberflächenwasser mit einer Pütz entnommen.
Die Phytoplanktonbiomasse wurde durch mikroskopische Zählung (UTERMÖHL-Methode) und die Chlorophyll-a-Konzentration durch Ethanol-Extraktion und fluorometrische Messung (Berechnung ohne Phaeopigment-Korrektur) bestimmt. Dabei wurden die Methodenvorschriften von HELCOM zugrundegelegt ( http://www.helcom.fi/Monas/CombineManual2/CombineHome.htm).
Die Zählung der Phytoplankton-Proben erfolgte bis zum 17.6.2008 mit dem Zählprogramm PhytoWin und ab dem 26.6.2008 mit OrgaCount. Die Kontinuität der Datenreihe wurde durch die Umstellung nicht gestört, da beide Programme auf der HELCOM-Artenliste (Olenina et al. 2006) basieren, die im April 2008 von der HELCOM-Phytoplankton-Expertengruppe aktualisiert wurde. Die spezifischen analytischen Bedingungen der Chlorophyll-a-Bestimmung sind in Wasmund et al. (2006) dargelegt. Es werden hier entsprechend der Entscheidung der BLMP-Unter-Arbeitsgruppe Qualitätssicherung vom 11.9.2008 die Chlorophyll-a-Konzentrationen angegeben, die nicht für Phaeopigment korrigiert sind.
An 12 Messtagen war es sturmbedingt zu hohen Sedimentanteilen in den Proben gekommen, die eine mikroskopische Auswertung unmöglich machten. Eine Chlorophyllbestimmung war aber möglich. Nur am 2.12.2008 fiel die Probennahme wegen Sturm komplett aus.
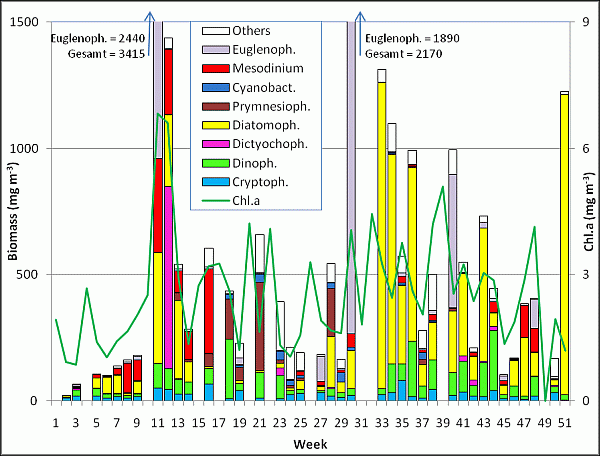
Erwartungsgemäß war die Phytoplankton-Biomasse in den ersten Wochen des Jahres sehr gering. Sie setzte sich am 8.1.08 hauptsächlich aus Cryptophyceen (Plagioselmis prolonga, Teleaulax spp., Hemiselmis spp.) zusammen. Ab der 3. Woche (15.1.08) kamen Gymnodiniales und ab der 5. Woche (29.1.08) Ceratium tripos und Porosira glacialis hinzu. Erstaunlich ist der schnelle Wechsel innerhalb der Kieselalgen von Porosira glacialis zu Rhizosolenia setigera von der 5. zur 6. Woche und dann wieder zurück zu Porosira glacialis in der 7. Woche. Offensichtlich sind hier Wasserkörper mit unterschiedlicher Artensignatur an der Station vorbeigedriftet. Ab der 7. Woche (12.2.08) entwickelte sich auch Mesodinium rubrum deutlich und erreichte in der 8.Woche eine Biomasse von 118 mg m-3. Am 26.2.08 trugen Mesodinium rubrum, Porosira glacialis, Eutreptiella sp., Teleaulax spp. und Guinardia delicatula jeweils mehr als 10 mg m-3 bei. Leider konnte die Probe vom 4.3.08 nicht ausgewertet werden, so dass eine genaue Angabe zum Beginn der Frühjahrsblüte nicht möglich ist.
Am 11.3.08 hat die Frühjahrsblüte bereits ihr Maximum (3415 mg m-3) erreicht. Sie wurde erstaunlicherweise nicht wie üblich von Kieselalgen dominiert, sondern von der Euglenophycee Eutreptiella spp. (2440 mg m-3). Darüber hinaus waren die Kieselalgen Rhizosolenia setigera und Rhizosolenia hebetata formasemispina, sowie der autotrophe Ciliat Mesodinium rubrum bedeutsam. Auch Teleaulax spp., Gyrodinium spirale, Protoperidinium depressum, Ceratium tripos und Proboscia alata sind erwähnenswert.
Die Frühjahrsblüte klang schnell wieder ab, wobei auch hier ein Transport von Wasserkörpern stattgefunden haben muss, denn am 18.3.08 (12.Woche) unterschied sich die Artenzusammensetzung grundlegend von der des 11.3.08 (11. Woche). Eutreptiella spp. fehlte fast völlig und es trat statt dessen der zuvor nicht vorhandene Kieselflagellat Dictyocha speculum mit 722 mg m-3 auf. Da er meist in seiner nackten Form vorkam, ist die Bestimmung nicht sicher; es könnten auch die Gattungen Chattonella, Verrucophora oder Pseudochattonella in Frage kommen. Unter den Kieselalgen dominierten Rhizosolenia hebetata forma semispina, Proboscia alata, Rhizosolenia setigera und Guinardia delicatula. Ein ungewöhnlich frühes Auftreten des Maximums von Dictyocha speculum wurde auch im Jahre 2007 beobachtet. Normalerweise ist die Dictyocha-Blüte erst nach der Kieselalgenblüte im April zu erwarten. Sie verdrängte im Jahre 2007 jedoch die Kieselalgen und auch Dinoflagellaten konnten sich nur gering entwickeln (Wasmund et al. 2008). Die ungewöhnliche Entwicklung des Jahres 2007 wiederholte sich also im Jahre 2008 und könnte sich möglicherweise dauerhaft etablieren (Regime Shift ?).
Zum 25.3.08 (13. Woche) waren sowohl Eutreptiella spp. als auch Dictyocha speculum fast vollständig verschwunden. Nun war plötzlich das zuvor unbedeutende Skeletonema costatum mit 270 mg m-3 dominant. Es ist regelmäßig eine der bedeutendsten Frühjahrs-Kieselalgen. Im Jahre 2008 wurde eine Frühjahrsblüte der Kieselalgen aber nicht gefunden. Trotz erforderlicher Neubewertung der Skeletonema-Arten (vgl. Sarno et al., 2005) behalten wir vorläufig den Namen Skeletonema costatum bei.
Zum 15.4.08 (16. Woche) waren die Kieselalgen vollständig verschwunden und es traten nun vermehrt Organismen auf, die sich mixotroph ernähren könnten, wie Cryptophyceen (Plagioselmis prolonga) und Prymnesiophyceen (Chrysochromulina spp.). Eventuell gehört auch der nun dominierende Ciliat Mesodinium rubrum in diese Gruppe.
Zum 29.4.08 (18. Woche) fand wiederum ein Wechsel statt: Mesodinium rubrum und Cryptophyceen waren nicht mehr vorhanden, während sich Chrysochromulina spp. verstärkte. Dinoflagellaten (Heterocapsa triquetra, Heterocapsa rotundata) wurden dominant. Das sehr frühe Auftreten von Aphanizomenon sp. (12 mg m-3) ist erwähnenswert. Am 20.5.08 (21. Woche) war die Phytoplankton-Zusammensetzung ähnlich.
Zum 3.6.08 (23. Woche) war Chrysochromulina spp. fast vollständig verschwunden. Es dominierten nun kleine unbestimmte Einzeller. Unter ihnen könnten sich durchaus noch schwer identifizierbare Chrysochromulina spp. befinden, ansonsten aber wohl hauptsächlich coccale Cyanobakterien.
Zum Ende des Juni nimmt die Biomasse weiter ab. Kaum eine Art erreicht nun eine Biomasse von über 10 mg m-3. Die dominierenden „Unbestimmten“ werden meistens durch kleine coccale Cyanobakterien (< 2 µm) und sehr dünne fädige Cyanobakterien (< 1 µm) gebildet. Am 8.7.08 (Woche 28) waren plötzlich Chrysochromulina sp. und Proboscia alata dominant. Letztere war auch noch am 22.7.08 (Woche 30) die dominierende Kieselalge. Hervorzuheben ist allerdings die extreme spontane Auftreten des Flagellaten Eutreptiella sp., das nicht von einem entsprechenden Chlorophyll-Signal begleitet wurde.
Die Proben vom 12.8. bis 2.9.08 (Woche 33-36) waren durch hohe Kieselalgenbiomassen gekennzeichnet, wobei in diesem Zeitraum eine Sukzession von Dactyliosolen fragilissimus zu Coscinodiscus granii, Guinardia flaccida und zur Pseudo-nitzschia seriata-Gruppe zu verzeichnen war. Langsam kam es auch zur üblichen Vermehrung von Ceratium tripos. In Woche 40 waren die unbestimmten Einzeller relativ groß (> 2 µm) und wurden deshalb mitgezählt, während vor und nach diesem Termin ebenfalls unbestimmte Einzeller auftraten, die wegen ihrer Größe von < 2 µm nicht gezählt wurden.
Am 7.10.08 (Woche 41) traten hauptsächlich Cerataulina pelagica, Coscinodiscus granii und Pseudo-nitzschia pungens auf. Letztere entwickelte bis zum 21.10.08 (Woche 43) eine Biomasse von 448 mg m-3. Am 28.10.08 dominierten die Dinoflagellaten Ceratium tripos, C. lineatum und C. fusus und am 11.-18.11.08 (Woche 46-47) plötzlich die Kieselalge Cerataulina pelagica. Auch Mesodinium rubrum wurde wieder bedeutsam.
Erstaunlich war das späte Auftreten einer Herbstblüte von Coscinodiscus granii erst am 16.12.08 (Woche 51). Wir hätten sie bei Ausstrom aus der Ostsee erwartet, aber der relativ hohe Salzgehalt von 13.3 PSU spricht eher für Einstrom oder upwelling.
Es zeigte sich wiederum, dass die Küstenstation für eine Aufzeichnung der Phytoplanktonsukzession nicht optimal ist. Die großräumige Algenentwicklung (z.B. Zeitpunkt der Blüte, Höhe der Biomasse) konnte zwar gut widergespeigelt werden. Die abrupten Änderungen der Artenzusammensetzung in der Datenserie, können allerdings nicht im Sinne einer Sukzession gewertet werden. Sie beruhen sicher auf großräumigen Unterschieden in der Artenzusammensetzung, so dass das Vorbeidriften der verschiedenen Wasserkörper an der Untersuchungsstation zu diesen Artenveränderungen führt, die nicht mit zeitlichen Entwicklungen zu erklären sind. Eine Kombination mit Monitoringdaten aus den offenen Seegewässern und mit Strömungsdaten, würde auch die Beschreibung räumlicher Heterogenitäten ermöglichen. Die Daten des HELCOM-Monitoring stehen allerdings erst mit starker zeitlicher Verzögerung zur Verfügung.
Literatur:
Olenina, I., Hajdu, S., Andersson, A.,Edler, L., Wasmund,N., Busch, S., Göbel, J., Gromisz, S., Huseby, S., Huttunen, M., Jaanus, A., Kokkonen, P., Ledaine, I., Niemkiewicz, E. (2006): Biovolumes and size-classes of phytoplankton in the Baltic Sea. Baltic Sea Environment Proceedings No.106, 144pp.
Internet-Zugang:
Paper: http://www.helcom.fi/stc/files/Publications/Proceedings/bsep106.pdf
Tabelle: http://www.helcom.fi/stc/files/Publications/Proceedings/bsep106ANNEX1Biovolumes_web.xls
Sarno, D., Kooistra, W.H.C.F., Medlin, L., Percopo, I., Zingone, A. (2005). Diversity in the genus Skeletonema (Bacillariophyceae). II. An assessment of the taxonomy of S. costatum-like species with the description of four new species. J. Phycol. 41, 151-176.
Wasmund, N., Topp, I., Schories, D. (2006): Optimising the storage and extraction of chlorophyll samples. Oceanologia 48: 125-144.
Wasmund, N., Pollehne, F.; Postel, L., Siegel, H., Zettler, M.L. (2008): Biologische Zustandseinschätzung der Ostsee im Jahre 2007. Meereswiss. Ber., Warnemünde, 74, 88 pp.
IOW, 29.01.2009
Dr. Norbert Wasmund,
Susanne Busch,
Ina-Marie Topp,
Regina Hansen.

Der Zustand der Ostsee
- Jährliche Zustandseinschätzungen
- Fahrtberichte
- Daten der autonomen Messstationen
- Faktenblätter
- Kontinuierliche CO2- und O2-Messungen auf Frachtschiff FINNMAID
- Entwicklung der suboxischen und anoxischen Bereiche seit 1969
- Baltic Thalweg Transekte seit 2014
- Algenblüten vor Heiligendamm seit 1998
- Salzwassereinbruch Dezember 2014
- Salzwassereinbruch Januar 2003
- Barotrope Salzwassereinbrüche 1887 - 2018
- Weiterführende Literatur

IOW-Fotogalerie einzelliger Ostseealgen
... licht- und elektronenmikroskopische Aufnahmen pelagischer Mikroalgen