

Phytoplankton development at the coastal station "Seebrücke Heiligendamm" in 2012
The Leibniz Institute for Baltic Sea Research conducts a coastal monitoring programme with weekly samplings at the sea-bridge Heiligendamm (54°08,55' N; 11°50,60' E; 300 m off shore, 3 m water depth). The Department of Marine Biology analyses the surface samples, taken by means of a bucket, for phytoplankton composition and biomass and for chlorophyll a.
The phytoplankton biomass is determined by microscopical counting (UTERMÖHL method) and the chlorophyll a concentration by ethanol extraction and fluorometric measurement. Method instructions see http://www.helcom.fi/groups/monas/CombineManual/AnnexesC/en_GB/. Phytoplankton counting was carried through by use of the counting programme OrgaCount and is based on the HELCOM-biovolume factors which are annually updated: http://www.ices.dk/env/repfor/index.asp (PEG_BVOL2012; basics see in Olenina et al. 2006). The analytical specifics of the chlorophyll a determination are published by Wasmund et al. (2006). According to the decision of the BLMP-subgroup “Quality Assurance” from 11.9.2008 we show here chlorophyll a data which are not corrected for pheopigments.
Sampling was not possible on 24.07.2012 because of reconstruction of the sea-bridge. Moreover, microscopical analysis was not possible in some samples if they contained high sediment portions, caused by wind-induced sediment resuspension at the shallow station. It seems that the share of sediment-containing samples increased, probably because of diminishing water depth.
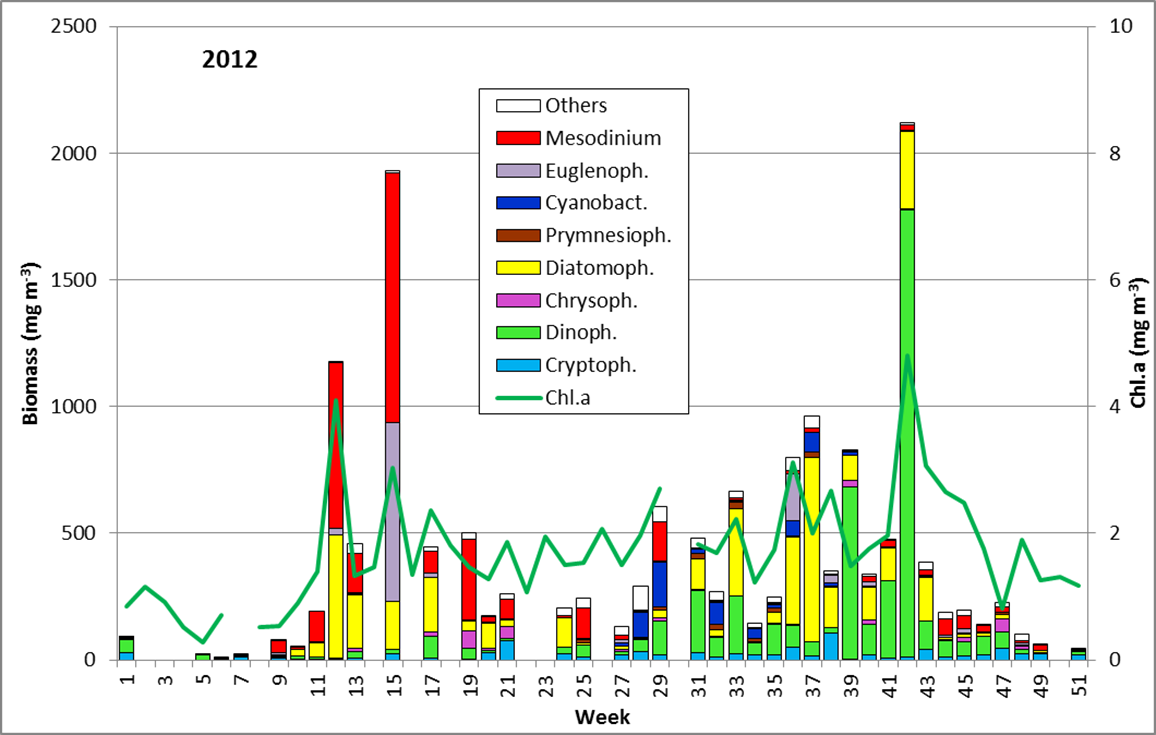
The results are shown in Fig. 1
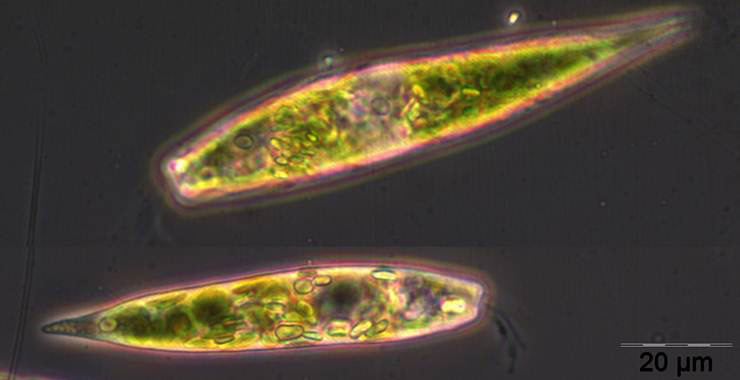
Despite gaps in the phytoplankton data series, we think that phytoplankton biomass was generally low in January and February 2012, as suggested by low chlorophyll-a-concentration <1 mg m-3. In January, Ceratium tripos and Teleaulax sp. were dominant. In February, Mesodinium rubrum developed, and since 6.3.2012 (week 10) also diatoms (Diatomophyceae). Mesodinium rubrum and diatoms reached a first maximum by the 20.3.2012 (week 12). Also the Euglenophyceae Eutreptiella braarudii (Image 1) appeared, like in the previous year.
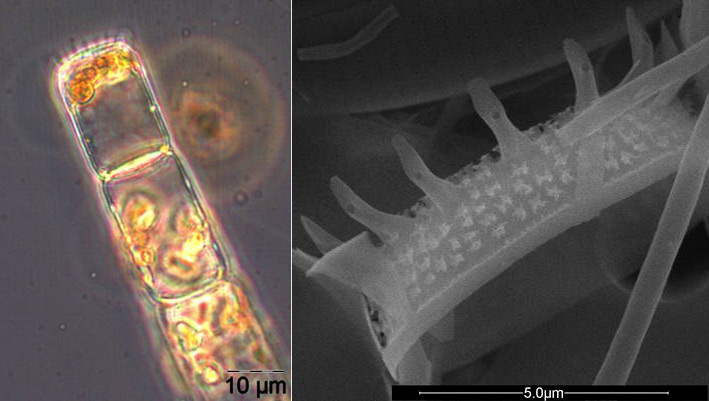
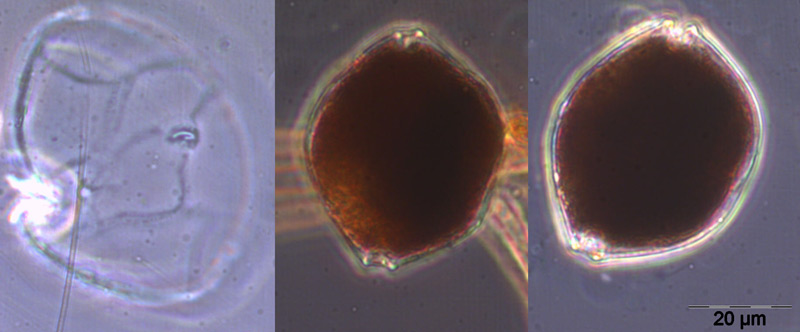
On the day of this spring diatom maximum, Skeletonema sp. (Image 2), Detonula confervacea (Image 3), Chaetoceros spp. (mainly Ch. wighamii) and Rhizosolenia setigera, were the dominating diatoms, but not Achnanthes taeniata anymore. The spring diatom maximum appeared at the same time as in the previous year, but was considerably lower. In comparison with 2011, Mesodinium rubrum was much more abundant, but Dictyocha speculum was almost completely absent in spring 2012.
After a significant biomass decrease, a second spring maximum appeared on 10.4.2012 (week 15) which contained less diatoms (Skeletonema sp.; image 2.) but relatively high biomass of Eutreptiella sp., a flagellate which was important already in 2008 and 2010. That species disappeared rather quickly. Despite lacking samples from before and after the 10.4.2012, chlorophyll-data show that this bloom was only of short duration.

In general, the spring bloom, especially the diatoms, was much lower in 2012 than in the previous year. Dictyocha speculum (in Fig. 1 counted to the Chrysophyceae), which was an abundant spring species in some years, appeared only on 8.5. and 22.5.2012 (week 19 and 21) in rather low abundances. Also the frequently recorded shift from diatoms to dinoflagellates during the spring was not noticed in 2012.
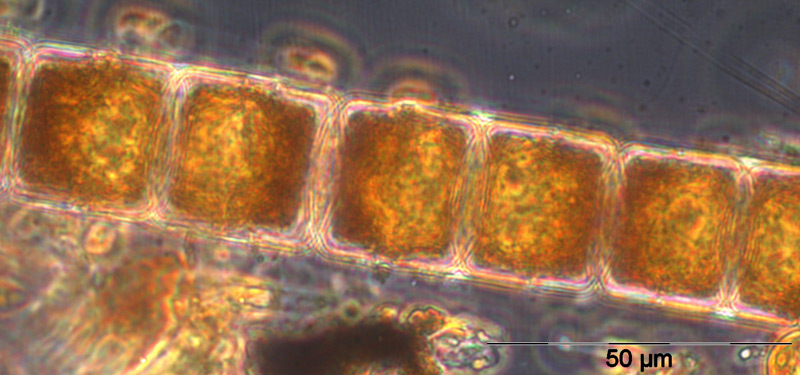
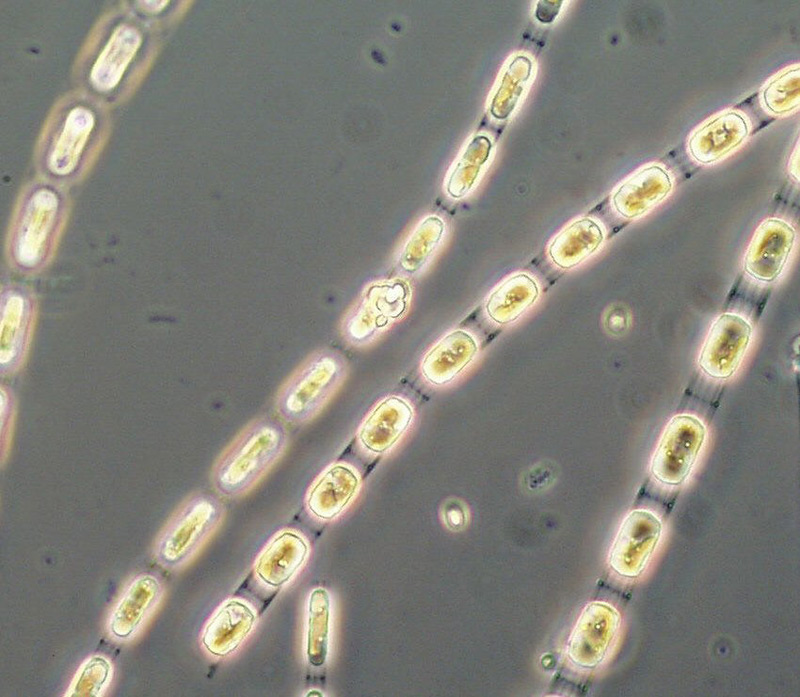
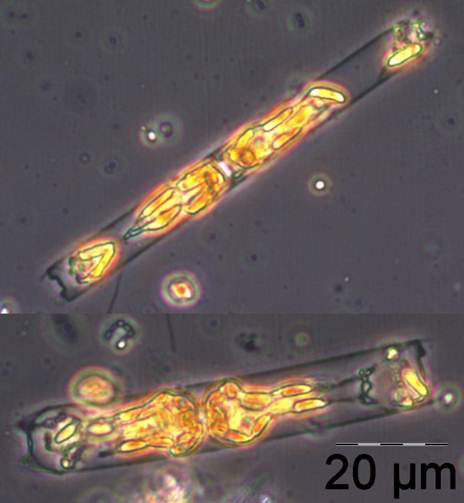
The appearance of the large diatom Coscinodiscus radiatus (image 4) is unusual in spring; it reached 82 mg m-3 on 15.5.2012 (week 20). In the following weeks, phytoplankton biomass stayed low, but diversity was high. On 12.6.2012 (week 24), Melosira moniliformis (image 5) dominated besides Dactyliosolen fragilissimus, Heterocapsa rotundata and Plagioselmis prolonga, and the development of Ceratium tripos started. Mesodinium rubrum appeared again on19.6.2012, besides a lot of heterotrophic ciliates (Strombidium). On this day, the nitrogen-fixing cyanobacteria Anabaena spp.and Aphanizomenon sp. emerged, which stayed present, beside the colony-forming Cyanobacteria, like Aphanocapsa spp. and Snowella spp. until autumn. Since the 10.7.2012, Nodularia spumigena (image 6) grew to 134 mg m-3 by the 17.7.2012 (week 29), disappeared shortly, reappeared on 7.8.2012 with 61 mg m-3 and stayed present with reducing biomass until the end of September.
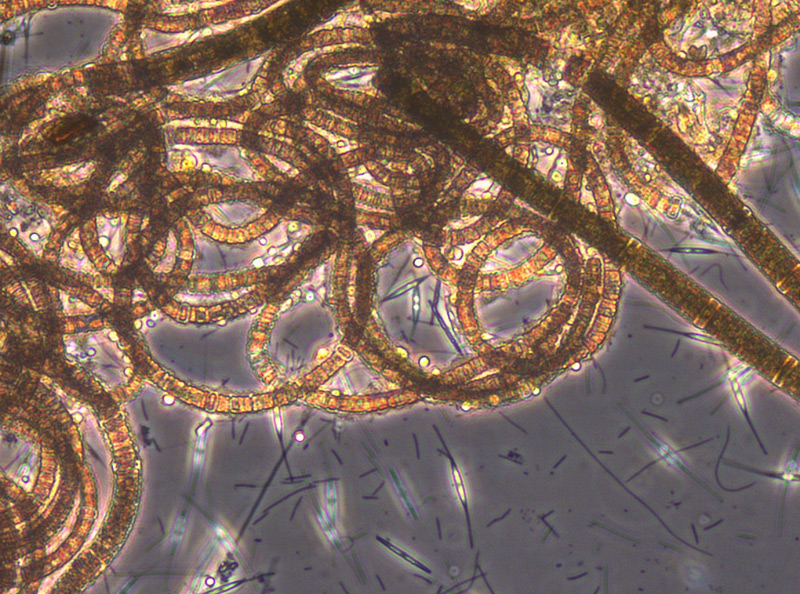
Besides Mesodinium rubrum, also Cladopyxis claytonii and Cymbomonas tetramitiformis are worth mentioning on 17.7.2012. The dinoflagellate Alexandrium pseudogonyaulax (Image 7) reached 137 mg/m³ on 31.7.2012 (week 31) and was still more important than in the year 2011.
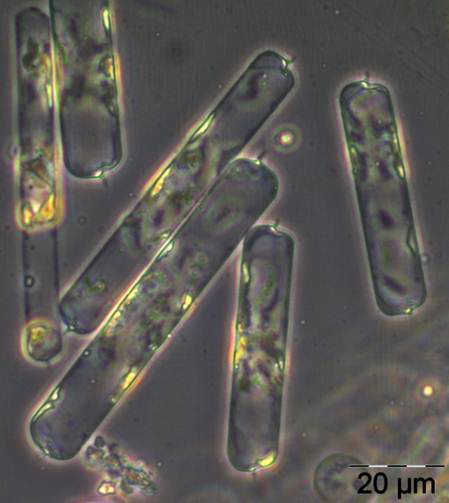
In the following weeks, Ceratium tripos stays important, but also other dinoflagellates (e.g. Prorocentrum micans) and typical summer-diatoms, such as Proboscia alata, Cerataulina pelagica (image 8) and Dactyliosolen fragilissimus (image 9) develop. These diatoms decreased strongly by the end of September (Week 39), while Ceratium-species (C. tripos, C. lineatum, C. fusus; see image 10) grew strongly. Ceratium-species reached their maximum on 9.10.2012 (week 42), when Ceratium tripos alone accounted for 1120 mg m-3.
On this day, also Pseudo-nitzschia pungens, Guinardia flaccida, Prorocentrum micans, Protoperidinium divergens, Dinophysis norvegica, Chaetoceros affinis, Ditylum brightwellii and Mesodinium rubrum are worth mentioning.
Pseudosolenia calcar-avis, noticed by us for the first time in 2010 and forming a strong bloom in September 2011, was unimportant in 2012. Also a bloom of Coscinodiscus granii , that was recorded in some years, was not observed in 2012. This species accounted for only 66 mg m-3 at its maximum on 23.10.2012 (week 43). Since the 30.10.2012 (Week 44), phytoplankton biomass is low and dominated by Ceratium tripos and Mesodinium rubrum, temporarily also by Dictyocha speculum.

A prolongation of the growth period into the winter, as discussed in the report of 2010, could not be found in 2012. In general, the diatom presence was low in 2012.
References:
Olenina, I., Hajdu, S., Andersson, A.,Edler, L., Wasmund, N., Busch, S., Göbel, J., Gromisz, S., Huseby, S., Huttunen, M., Jaanus, A., Kokkonen, P., Ledaine, I., Niemkiewicz, E. (2006): Biovolumes and size-classes of phytoplankton in the Baltic Sea. Baltic Sea Environment Proceedings No.106, 144pp.
Internet:
Paper: http://www.helcom.fi/stc/files/Publications/Proceedings/bsep106.pdf
Tabelle: http://www.helcom.fi/stc/files/Publications/Proceedings/bsep106ANNEX1Biovolumes_web.xls
Wasmund, N., Topp, I., Schories, D. (2006): Optimising the storage and extraction of chlorophyll samples. Oceanologia 48: 125-144.
IOW, 17.01.2013
Dr. Norbert Wasmund,
Susanne Busch,
Christian Burmeister,
Regina Hansen.
Leibniz Institute for Baltic Sea Research Warnemünde (IOW),
Seestr. 15,
D-18119 Rostock-Warnemünde
Corresponding author: Dr. Norbert Wasmund

State of the Baltic Sea
- Annual Reports on the state of the Baltic Sea Environment
- Cruise Reports
- Data from the autonomous measuring stations
- Development of the suboxic and anoxic regions since 1969
- Baltic Thalweg transect since 2014
- Algal blooms at Heiligendamm since 1998
- "Major Baltic Inflow" December 2014
- "Major Baltic Inflow" January 2003
- Baltic saline barotropic inflows 1887 - 2018
- Further Reading
