

Phytoplankton development at the coastal station “Seebrücke Heiligendamm” in 2018
The departments of Marine Chemistry and of Biological Oceanography of the Leibniz Institute for Baltic Sea Research conduct a coastal monitoring programme with weekly samplings at the jetty Heiligendamm (54°08,76' N; 11°50,58' E; nearly 3 m water depth). This report presents the results of the investigations on phytoplankton composition and biomass and on chlorophyll a.
Surface samples were taken by means of a bucket. The phytoplankton biomass is determined by microscopic counting (UTERMÖHL method) and the chlorophyll a concentration by ethanol extraction and fluorometric measurement. Method instructions see:
http://www.helcom.fi/Documents/Action%20areas/Monitoring%20and%20assessment/Manuals%20and%20Guidelines/Guidelines%20for%20monitoring%20phytoplankton%20species%20composition,%20abundance%20and%20biomass.pdf
Phytoplankton counting was carried through by use of the counting programme OrgaCount and is based on the HELCOM-biovolume factors which are annually updated: http://www.ices.dk/marine-data/vocabularies/Documents/PEG_BVOL.zip (basics see Olenina et al. 2006). The analytical specifics of the chlorophyll a determination are published by Wasmund et al. (2006) and in the annual biological assessments of the Baltic Sea (e.g. Wasmund et al. 2018). The chlorophyll-a data are not corrected for pheopigments.
Microscopical analysis was not possible in 11 samples as they contained high sediment portions, caused by wind-induced sediment resuspension at the shallow station. The chlorophyll analyses were still possible and allowed information on phytoplankton biomass even if phytoplankton data were lacking. Samplings were not carried through on 27.2., 3.4. and 25.12.2018. Additional chlorophyll samples were taken in duplicate on 26.3.2018 in order to check and verify the extremely high chlorophyll values from 20.3.2018.
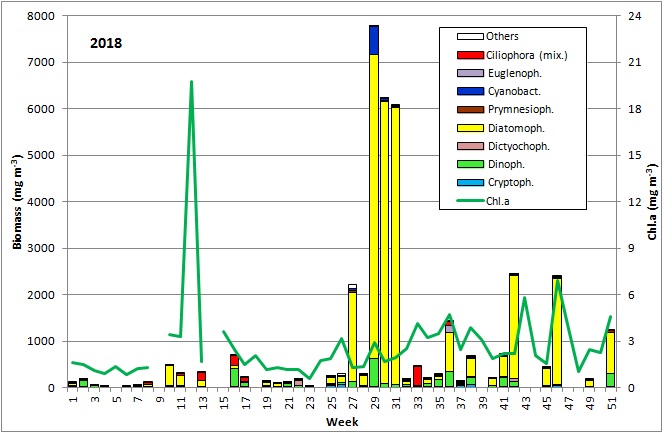
The results are shown in Fig. 1.
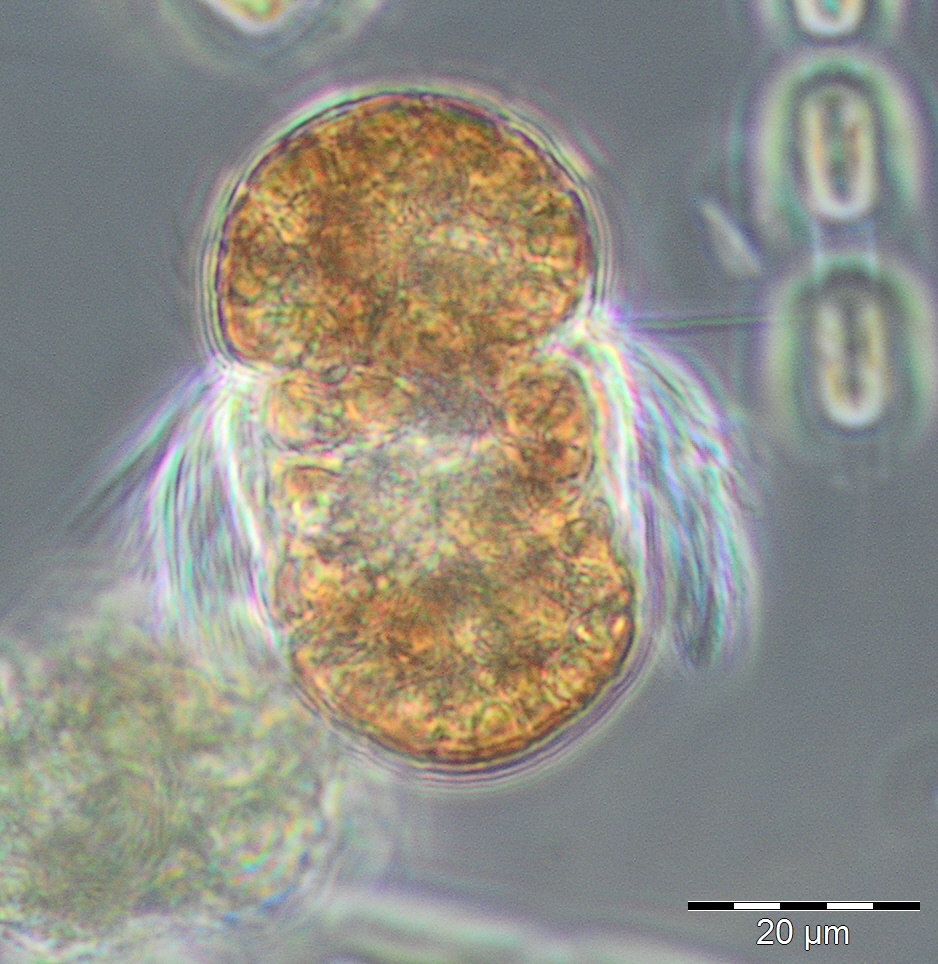
The first 8 weeks of the year 2018 were characterised by low phytoplankton biomass. However, they might have been underestimated because the samples contained much sediment that may have disturbed the identification of the full community. The relatively low chlorophyll concentrations confirm the low biomass values. Only in week 2, an elevated biomass of Ceratium tripos (105 mg/m3) was found. A scheduled sampling on 27.2.2018 (week 9) was cancelled because of a snow storm. On 6.3.2018 (week 10), chlorophyll concentrations and biomass were clearly increased and mark the start of the spring bloom. Diatoms (diatomophyceae) dominated with Skeletonema marinoi (160 mg/m3) and Thalassiosira species (194 mg/m3), with T. anguste-lineata being the most abundant in this genus (60 mg/m3). The mixotrophic ciliate Mesodinium rubrum (Image 1) appeared with 48 mg/m3 on 14.3.2018 (week 11).

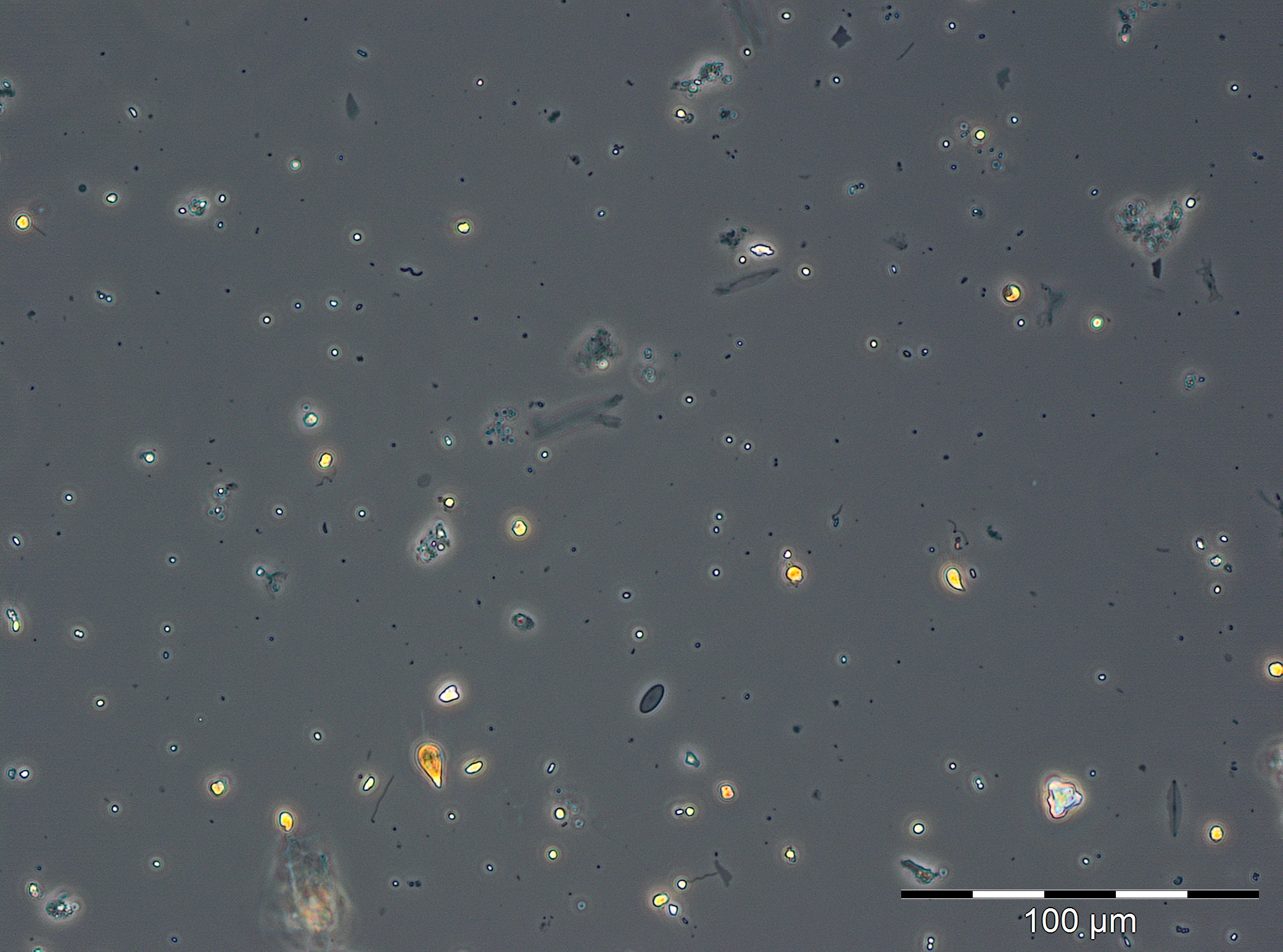
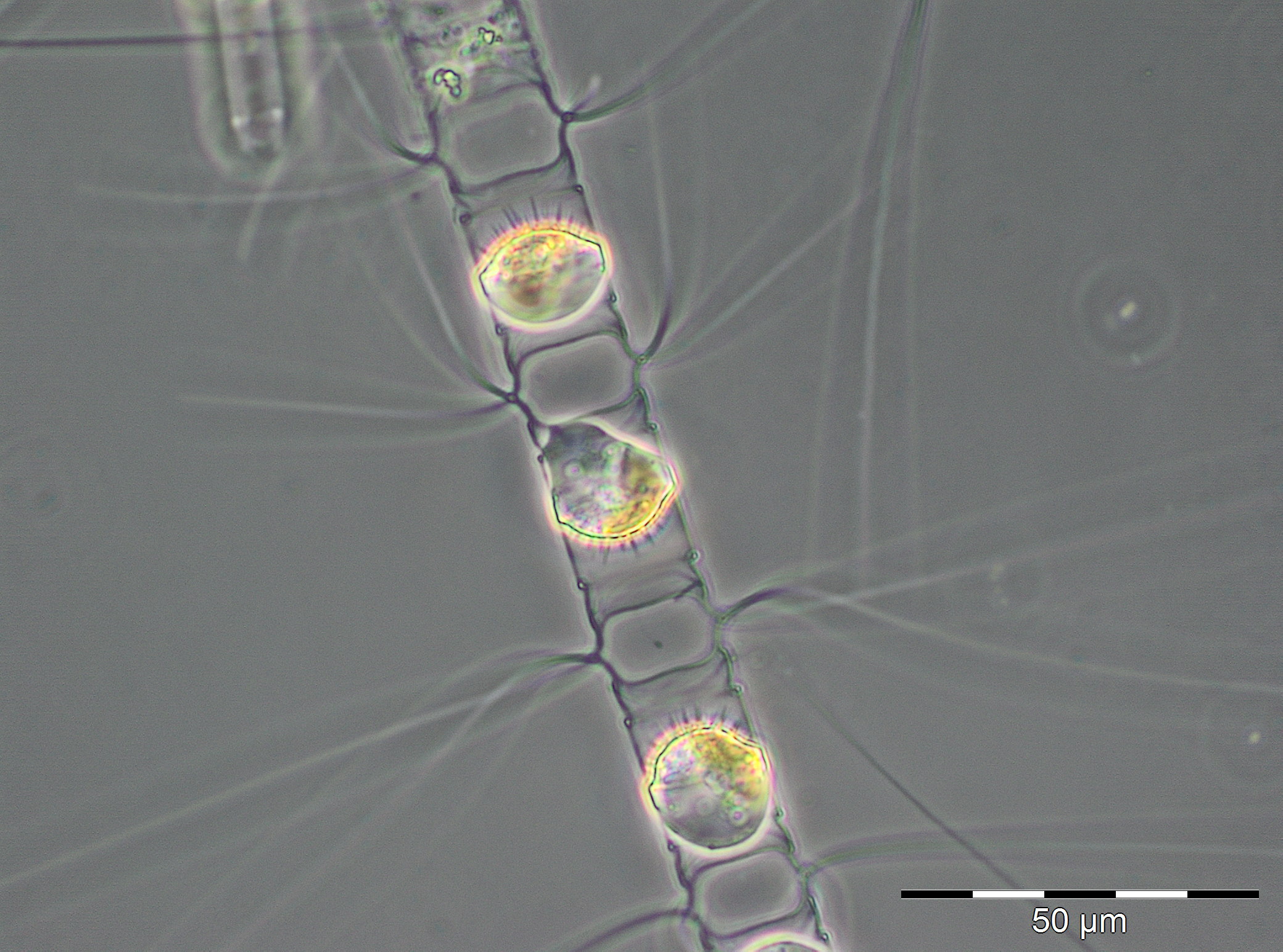
In the following week (20.3.2019), the chlorophyll concentration reached an unusually high peak value of 19.74 mg/m3. We could not verify it by phytoplankton biomass data because the corresponding phytoplankton sample could not be quantified due to a high sediment portion (Image 2). However, a high share of diatoms, especially Skeletonema marinoi and Thalassiosira species, could be detected. The high sediment share was caused resuspension due to strong north-easterly winds (6-8 Beaufort) on 20.3.2018 and may have caused also the high chlorophyll values. In order to verify the unusual chlorophyll data, an extraordinary chlorophyll sampling was conducted on 26.3.2019. The measurements (20.1 und 20.5 mg/m3) confirmed the data from 20.3.2018. Already on the next sampling occasion on 27.3.2019 (week 13), the bloom had collapsed or it was drifted away. Biomass and chlorophyll concentrations were low with 314 mg/m3 and 1.68 mg/m3, respectively. The phytoplankton was still dominated by Mesodinium rubrum, Skeletonema marinoi and Thalassiosira species.
A shift in phytoplankton composition occurred by 17.4.2018 (week 16). The diatoms have strongly declined and were represented mostly by Rhizosolenia setigera (51 mg/m3), but dinoflagellates have developed containing unidentified Gymnodiniales (294 mg/m3), Heterocapsa rotundata (39 mg/m3) and Peridiniella catenata (20 mg/m3). Mesodinium rubrum have increased (197 mg/m3). Rhizosolenia setigera and Peridiniella catenata have disappeared and Gymnodiniales as well as Mesodinium rubrum were reduced by 24.4.2018. This may mark the end of the spring bloom.
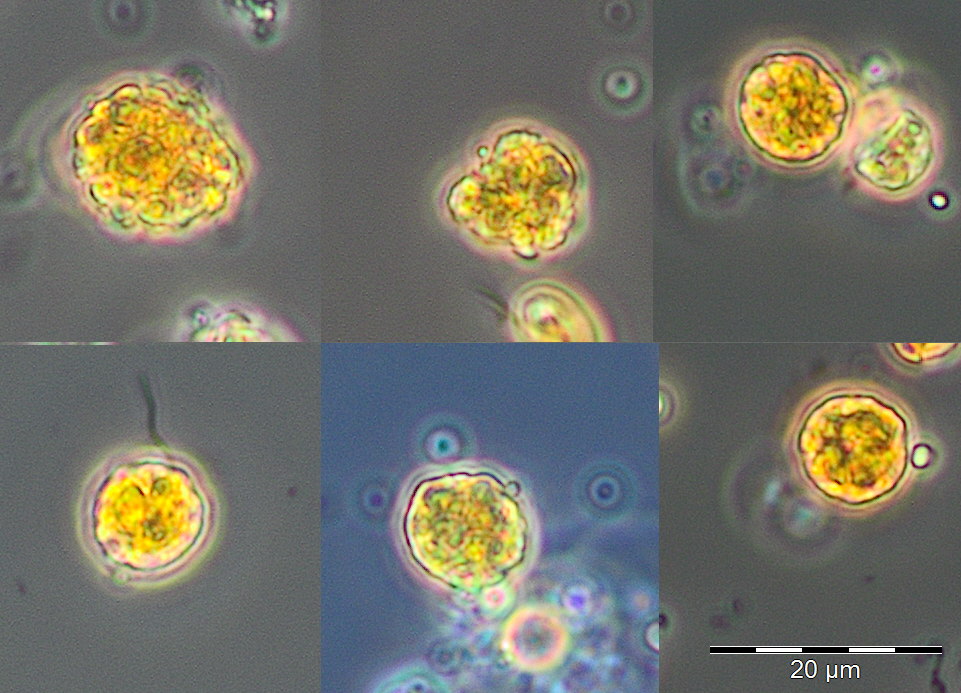
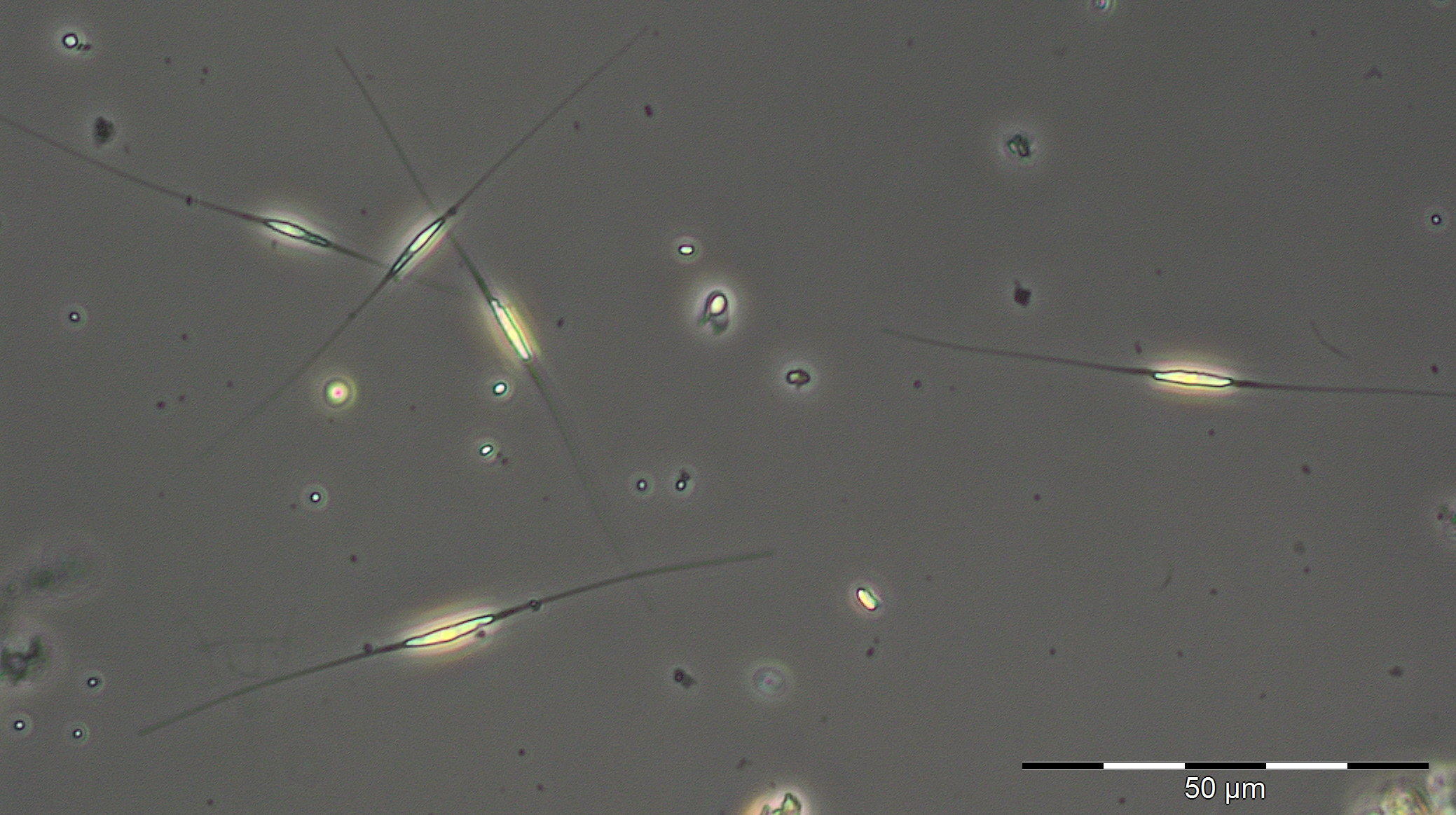
The period of the summer minimum of phytoplankton biomass started on 8.5.2019 (Week 19). Rhizosolenia setigera dominated on 8.5.2019 with 71 mg/m3. Also Plagioselmis prolonga, Hemiselmis sp. and Heterocapsa rotundata are worth mentioning, but Mesodinium rubrum has disappeared almost completely. An overview image from 22.5.2018 showing Cryptophyceae, Prymnesiales and unicells < 2µm (picoplankton) is presented as Image 3. On 30.5.2019 (week 22), Dictyocha speculum appeared shortly with 83 mg/m3 (Image 4). It formed large biomasses in late spring in some previous years (2008, 2011, 2014, 2015, 2017).
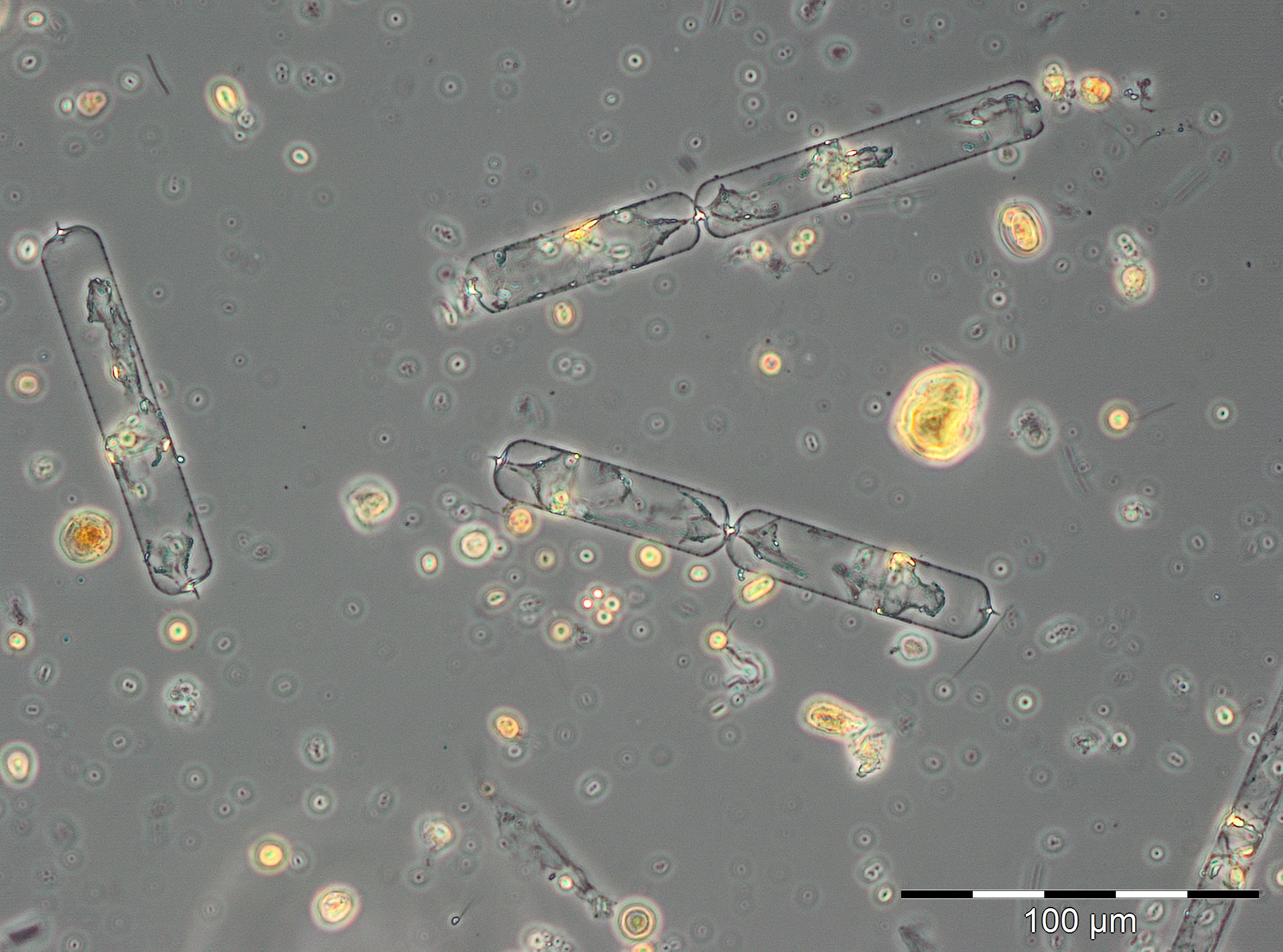
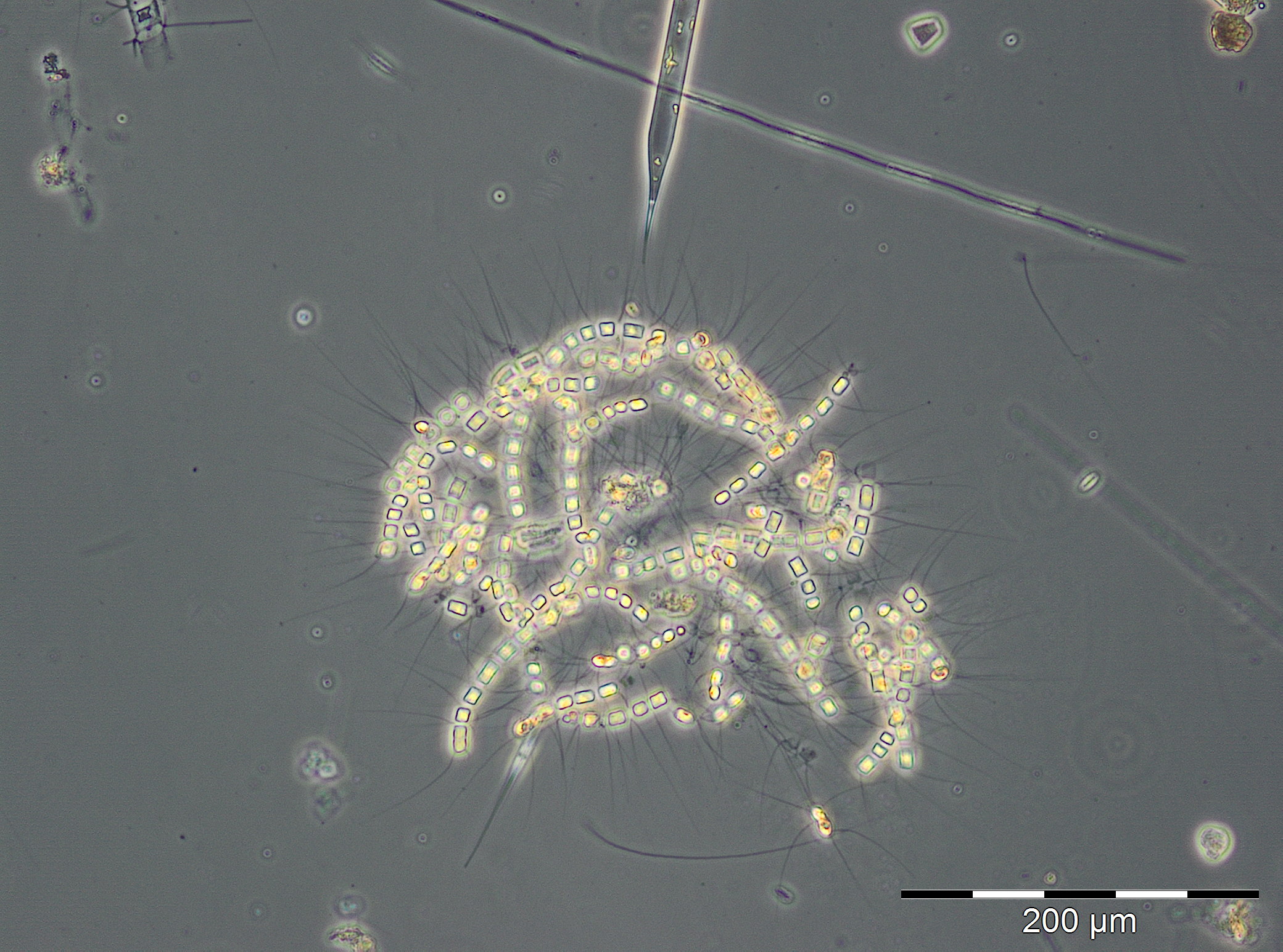
The summer bloom started on 19.6.2018 (week 25), but on this day it was unusually dominated by the spring species Skeletonema marinoi (120 mg/m3). This species disappeared almost completely already by the 26.6.2018, and it was replaced by the typical summer diatom Dactyliosolen fragilissimus (98 mg/m3), which has grown to 1916 mg/m3 on 3.7.2018 (Image 5). Such large diatom cells have relatively small cell plasm and correspondingly low chlorophyll content as seen from Image 5 (cf. Fig. 1). Important accompanying species were: Ceratium tripos, Nodularia spumigena, Cymbomonas tetramitiformis, Alexandrium pseudogonyaulax and unidentified Prymnesiales. The drastically reduced biomass in week 28 may have been caused by underestimation due to high sediment portion or by arrival of a deviant water body at the station. Anyway, the diatom bloom continues from 17.7. to 31.7.2018 (week 29-31) with biomass of Dactyliosolen fragilissimus of about 6000 mg/m3.
The appearance of high biomass of Nodularia spumigena (567 mg/m3) on 17.7.2018 is interesting because cyanobacteria blooms are of general public interest. Cyanobacteria biomasses exceeding 200 mg/m3 may be considered as blooms according to Wasmund (1997). The finding must not overrated because buoyant cyanobacteria mats may shortly be concentrated by wind and currents especially near the coast, but they can also be dispersed quickly. However, beaches on the Polish coast were temporally closed because of toxic cyanobacteria mats („Zeit online“ from 25.7.2018). The German outer coasts were not affected. On 17.7.2018, also dinoflagellates were important: Prorocentrum cordatum (216 mg/m3), Ceratium tripos (195 mg/m3) and Alexandrium pseudogonyaulax (178 mg/m3).

After the disappearance of the diatom summer bloom, Mesodinium rubrum appeared spontaneously with 405 mg/m3 on 14.8.2018 (week 33). The samples from weeks 34 and 35 were hardly quantifiable; they contained much sediment and much picoplankton. By 4.9.2018 (week 36), diatoms have developed; in this case not Dactyliosolen fragilissimus, but Pseudosolenia calcar-avis (316 mg/m3; Image 6), Chaetoceros species (C. contortus, C. circinalis, C. affinis, C. curvisetus, C. laciniosus, C. socialis; Images 6-8), Ditylum brightwellii, Cerataulina pelagica and Thalassiosira spp., and also dinophyceae (Polykrikos schwartzii, Dinophysis acuminata, Ceratium spp.), euglenophyceae (Eutreptiella) and Mesodinium rubrum.
The species composition changed again by 18.9.2018 (week 38). Besides a new appearance of the summer diatom Dactyliosolen fragilissimus (124 mg/m3) also the typical diatom of autumn, Coscinodiscus granii, appeared (216 mg/m3). Chaetoceros spp., Pseudosolenia calcar-avis, euglenophyceae, Mesodinium rubrum and the dinoflagellates mentioned above have vanished almost completely. The sample from 2.10.2018 (week 40) was silty and the biomass may be underestimated, but the dominance of Coscinodiscus granii could be recognised. This species disappeared by 9.10.2018 (week 41), when Pseudosolenia calcar-avis (219 mg/m3) dominated. P. calcar-avis has established since 2010. Its strong increment was also noted by Kaiser et al. (2016). Lennoxia faveolata was present in low biomass on 9.10.2018 (Image 9); it appears in our sample sporadically only since 2009 (cf. Wasmund et al. 2018).
Pseudosolenia calcar-avis was strongly reduced (78 mg/m3) by 16.10.2018 (week 42), but Cerataulina pelagica has grown extensively (1909 mg/m3). Worth mentioning is the presence of Dictyocha speculum in this sample (57 mg/m3) because this is primarily a spring species. The next evaluable sample (8.11.2018, week 45) showed again a completely different species composition with the following ranking order: Thalassiosira spp., Ditylum brightwellii, Pseudo-nitzschia spp., Proboscia alata, Pseudosolenia calcar-avis, Cerataulina pelagica. Obviously, a different water body was transported to the station. The sample from 13.11.2018 (week 46) fits again to the sample from week 42: Cerataulina pelagica was absolutely dominant (2129 mg/m3).

The following samples were not evaluable (Image 10). Also the data from week 49 shown in Fig. 1 may be an underestimate. The sample from 18.12.2018 (week 51) was unbiased. In this sample, Cerataulina pelagica (659 mg/m3) was still dominating, but other important species appeared: Pseudo-nitzschia spp. (202 mg/m3), Thalassiosira spp. (169 mg/m3), Ceratium tripos (140 mg/m3), Skeletonema marinoi (85 mg/m3), Mesodinium rubrum (44 mg/m3), Dactyliosolen fragilissimus (33 mg/m3).
In summary, the spring bloom 2018 was rather short (from about 6.3.2018 to 17.4.2018), as expected with a succession from diatoms (Skeletonema marinoi and Thalassiosira spp.) to motile species (dinoflagellates, Mesodinium rubrum). In contrast to some previous years, Dictyocha speculum was rare. A typical diatom summer bloom (Dactyliosolen fragilissimus) occurred from 3.7. to 31.7.2018 whereas a short local cyanobacteria bloom appeared on 17.7.2018. Since September, some smaller blooms of different composition were found, dominated by Pseudosolenia calcar-avis (4.9.2018, 9.10.18) and Cerataulina pelagica (16.10.-18.12.2018). The previously important autumn diatom Coscinodiscus granii and the typical dinoflagellate genus Ceratium showed relatively low biomass in 2018.
Acknowledgement
We wish to thank the colleagues of the department Marine Chemistry (Lars Kreuzer, Jenny Jeschek, Birgit Sadkowiak), who accompanied the joint sampling activities and analysed the nutrients, as well as the colleagues of the IT Group (Dr. Steffen Bock, Dr. Susanne Feistel, Solvey Hölzel), who processed the data for storage in the data base and loaded this report to the IOW homepage.
Literature:
Kaiser, J., Belt, S.T., Tomczak, M., Brown, T.A., Wasmund, N., Arz, H.W., 2016: C25 highly branched isoprenoid alkenes in the Baltic Sea produced by the marine planktonic diatom Pseudosolenia calcar-avis. – Organic Geochemistry 93: 51-58.
Olenina, I., Hajdu, S., Andersson, A.,Edler, L., Wasmund, N., Busch, S., Göbel, J., Gromisz, S., Huseby, S., Huttunen, M., Jaanus, A., Kokkonen, P., Ledaine, I., Niemkiewicz, E. (2006): Biovolumes and size-classes of phytoplankton in the Baltic Sea. Baltic Sea Environment Proceedings No.106, 144pp.
http://www.helcom.fi/Lists/Publications/BSEP106.pdf
Wasmund, N. (1997): Occurrence of cyanobacterial blooms in the Baltic Sea in relation to environmental conditions. Int. Revue ges. Hydrobiol. 82: 169-184.
Wasmund N., Dutz J., Pollehne F., Siegel H. & Zettler M.L. 2018. Biological assessment of the Baltic Sea 2017. Meereswiss. Ber., Warnemünde 108: 1-101.
https://www.io-warnemuende.de/files/forschung/meereswissenschaftliche-berichte/mebe108_2018_assessment-bio.pdf
Wasmund, N., Topp, I., Schories, D. (2006): Optimising the storage and extraction of chlorophyll samples. Oceanologia 48: 125-144.
Authors
IOW, 07.06.2019
Dr. Norbert Wasmund,
Susanne Busch,
Christian Burmeister.
Regina Hansen.
Leibniz Institute für Ostseeforschung Warnemünde (IOW),
Seestr. 15,
D-18119 Rostock-Warnemünde
Corresponding author: Dr. Norbert Wasmund

State of the Baltic Sea
- Annual Reports on the state of the Baltic Sea Environment
- Cruise Reports
- Data from the autonomous measuring stations
- Development of the suboxic and anoxic regions since 1969
- Baltic Thalweg transect since 2014
- Algal blooms at Heiligendamm since 1998
- "Major Baltic Inflow" December 2014
- "Major Baltic Inflow" January 2003
- Baltic saline barotropic inflows 1887 - 2018
- Further Reading
