

Phytoplanktonentwicklung an der Küstenstation Heiligendamm im Jahre 2018
Die Sektionen Biologische Meereskunde und Meereschemie des Leibniz-Instituts für Ostseeforschung Warnemünde (IOW) führen wöchentliche Probennahmen an der Seebrücke Heiligendamm (54°08,76' N; 11°50,58' E; etwa 3m Wassertiefe) durch. In diesem Bericht werden die Ergebnisse der Phytoplankton-Untersuchungen vorgestellt. Es wurde Oberflächenwasser mit einer Pütz entnommen.
Die Phytoplanktonbiomasse wurde durch mikroskopische Zählung (UTERMÖHL-Methode) und die Chlorophyll-a-Konzentration durch Ethanol-Extraktion und fluorometrische Messung bestimmt. Dabei wurden die Methodenvorschriften von HELCOM zugrunde gelegt: http://www.helcom.fi/Documents/Action%20areas/Monitoring%20and%20assessment/Manuals%20and%20Guidelines/Guidelines%20for%20monitoring%20phytoplankton%20species%20composition,%20abundance%20and%20biomass.pdf Die Zählung der Phytoplankton-Proben erfolgte mit dem Zählprogramm OrgaCount (AquaEcology) und basiert auf der HELCOM-Artenliste, die jährlich aktualisiert wird: http://www.ices.dk/marine-data/vocabularies/Documents/PEG_BVOL.zip (Grundlagen siehe in Olenina et al. 2006). Die spezifischen analytischen Bedingungen der Chlorophyll-a-Bestimmung sind in Wasmund et al. (2006) dargelegt sowie in den jährlichen Zustandseinschätzungen der Ostsee (Wasmund et al. 2018). Die Chlorophyll-a-Konzentrationen sind nicht für Phaeopigment korrigiert.
Die Phytoplanktonproben von 11 Messtagen konnten nicht mikroskopisch ausgewertet werden, da es sturmbedingt zu hohen Sedimentanteilen in den Proben gekommen war. Die Chlorophyllproben waren jedoch nicht betroffen, so dass sie Informationen über die Phytoplanktonbiomasse selbst bei fehlenden Phytoplanktonanalysen zuließen. Am 27.2., 3.4. und 25.12.2018 fielen die Probenahmen ganz aus. Am 26.3.2018 wurden zusätzliche Chlorophyll-Proben (Doppelbestimmung) genommen, um die extrem hohen Chlorophyll-Werte vom 20.3.2018 zu verifizieren.
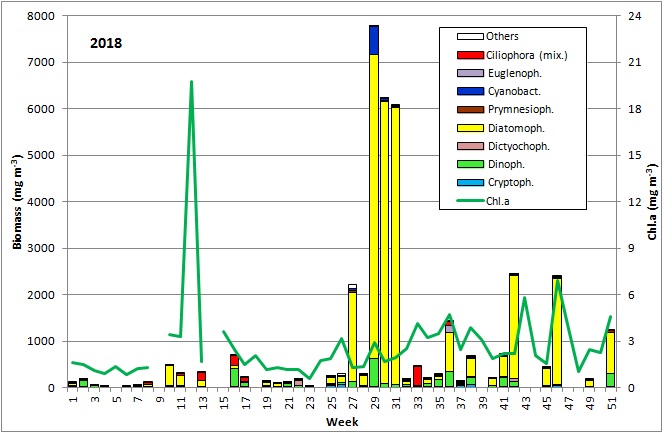
Die Ergebnisse sind in Abb. 1 dargestellt.
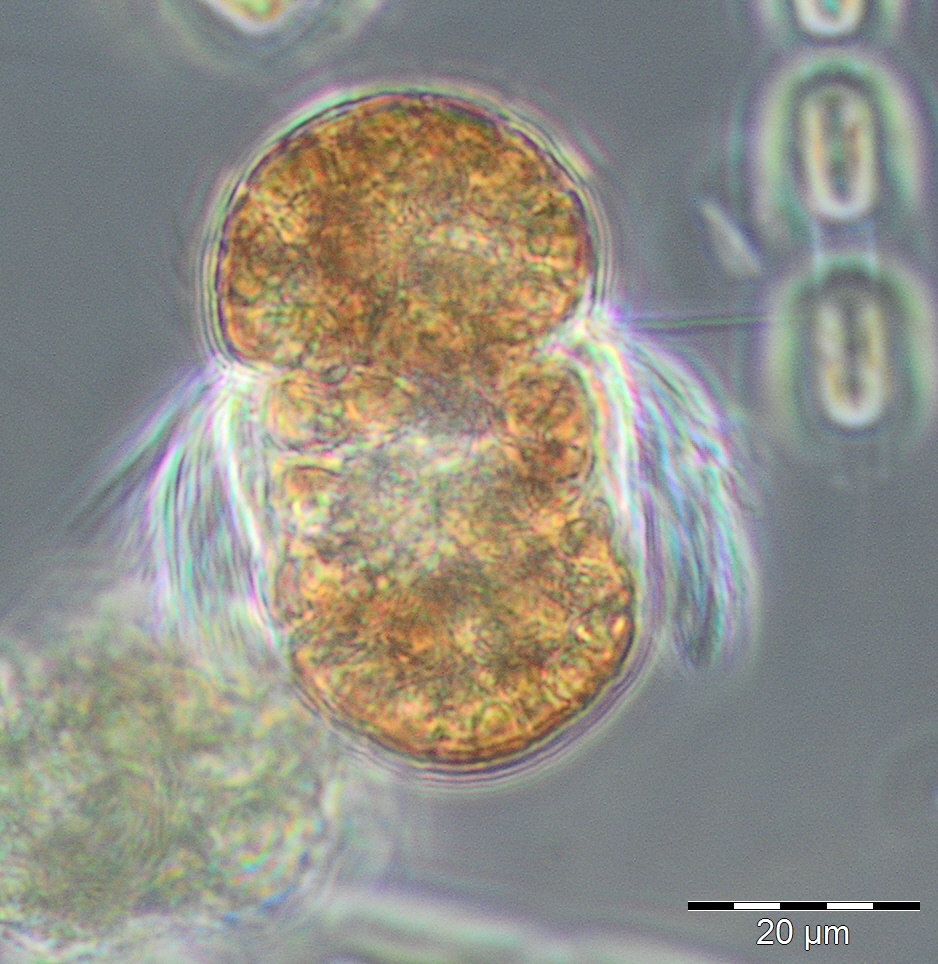
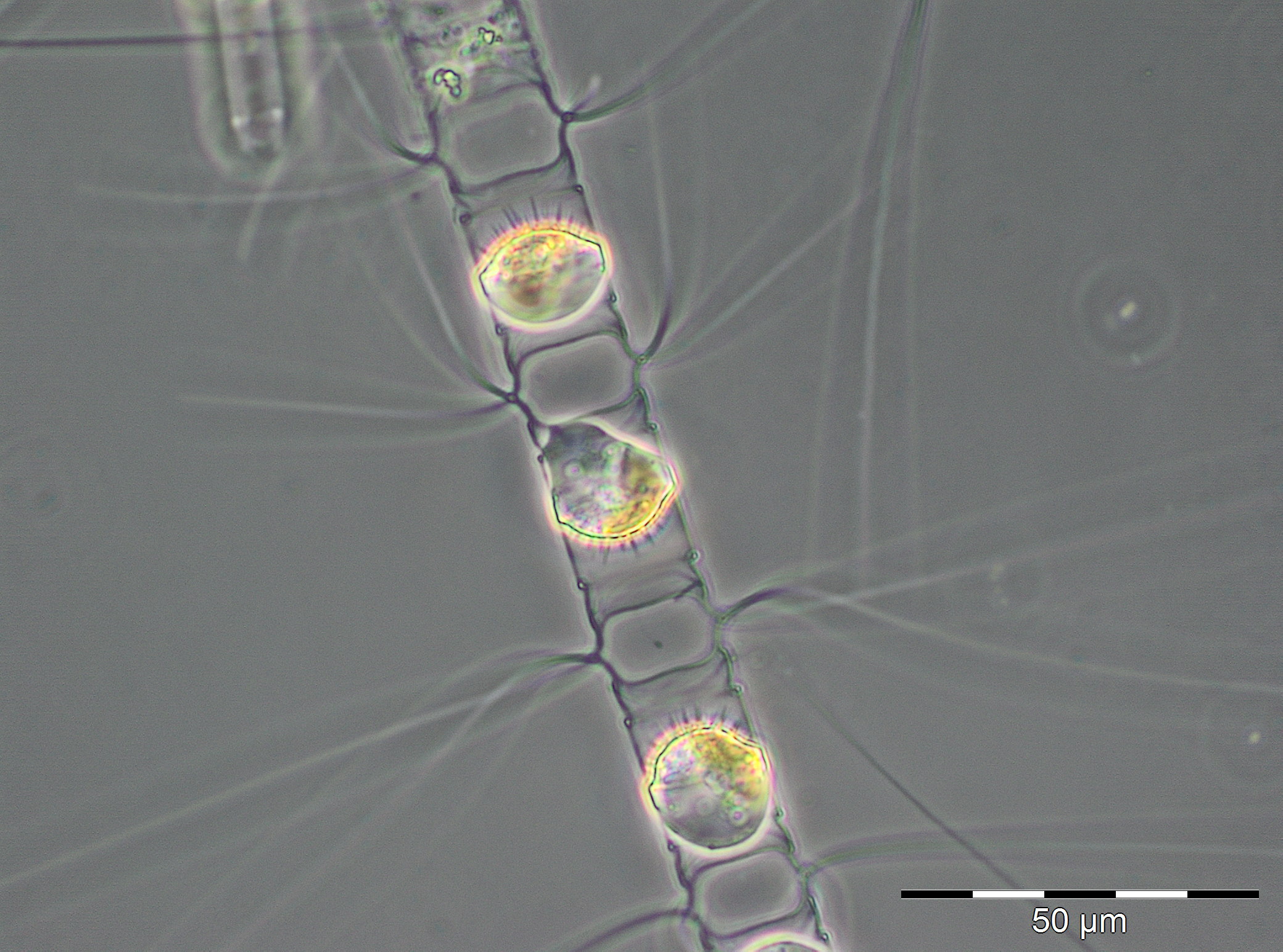
Die ersten 8 Wochen des Jahres waren durch geringe Biomassen gekennzeichnet, die allerdings wegen hohen Anteils an anorganischem Material und entsprechender schlechter Zählbarkeit der Proben unterschätzt sein könnten. Die relativ geringen Chlorophyll-Konzentrationen bestätigen jedoch die Biomasse-Daten. Lediglich in Woche 2 war eine erhöhte Biomasse durch Ceratium tripos (105 mg/m3) zu verzeichnen. Die Probenahme am 27.2.2018 (Woche 9) fiel wegen Schneesturms aus Sicherheitsgründen aus. Am 6.3.2018 (Woche 10) waren Chlorophyll-Konzentra-tion und Biomasse bereits deutlich erhöht und zeigen offensichtlich den Beginn der Frühjahrsblüte an. Es dominierten die Kieselalgen (Diatomophyceae) Skeletonema marinoi (160 mg/m3) und Thalassiosira-Arten (194 mg/m3), worunter T. anguste-lineata mit 60 mg/m3 die häufigste war. Am 14.3.2018 (Woche 11) kam der mixotrophe Ciliat Mesodinium rubrum hinzu (48 mg/m3, Bild 1).

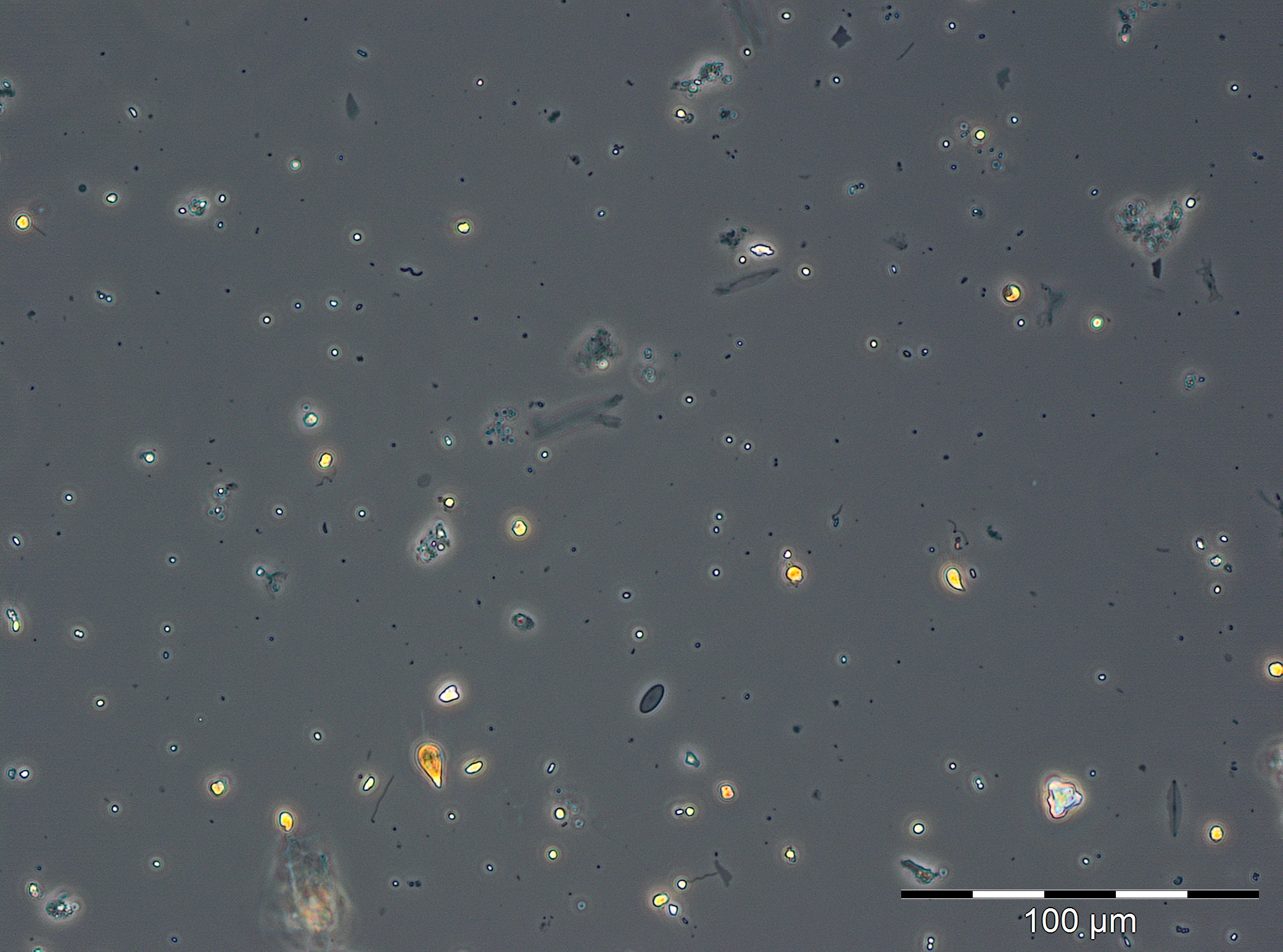
In der darauffolgenden Woche (20.3.2019) erreichte die Chlorophyll-Konzentration einen ungewöhnlich hohen Extremwert von 19,74 mg/m3. Dieser konnte nicht durch die Phytoplanktonprobe bestätigt werden, da diese wegen extrem starker Sedimentbeimischung nicht auswertbar war (Bild 2). Es zeigten sich in der Probe aber recht hohe Anteile an Kieselalgen, insbesondere Skeletonema marinoi und Thalassiosira-Arten. Die hohen Sedimentanteile waren durch starken Nordostwind (Windstärke 6-8) am 20.3.2018 bedingt und könnten auch zu dem hohen Chlorophyll-Wert beigetragen haben. Um zu prüfen, ob tatsächlich eine Blüte mit derart hohen Chlorophyll-Werten aufgetreten ist, wurden am 26.3.2019 zwei außerplanmäßige Chlorophyll-Proben genommen. Sie ergaben Chlorophyll-Konzentrationen von 20,1 und 20,5 mg/m3, bestätigten also den Wert vom 20.3.2018. Bereits zur regulären Probennahme vom 27.3.2019 (Woche 13) war die Phytoplanktonblüte zusammengebrochen oder verdriftet. Biomasse und Chlorophyll-Konzentration waren mit 314 mg/m3 bzw. 1,68 mg/m3 gering. Das Phytoplankton wurde immer noch durch Mesodinium rubrum, Skeletonema marinoi und Thalassiosira-Arten dominiert.
Bis zur nächsten verfügbaren Phytoplanktonprobe vom 17.4.2018 (Woche 16) hatte sich ein Wechsel der Phytoplankton-Zusammensetzung vollzogen. Die Kieselalgen war fast gänzlich verschwunden, im Wesentlichen vertreten durch Rhizosolenia setigera (51 mg/m3), aber Dinoflagellaten hatten sich entwickelt, darunter meistens unbestimmte Gymnodiniales (294 mg/m3), Heterocapsa rotundata (39 mg/m3) und Peridiniella catenata (20 mg/m3). Mesodinium rubrum hatte sich verstärkt (197 mg/m3). Zum 24.4.2018 waren Rhizosolenia setigera und Peridiniella catenata ganz verschwunden und Gymnodiniales sowie Mesodinium rubrum waren reduziert. Dieses dürfte das Ende der Frühjahrsblüte-Phase gewesen sein.
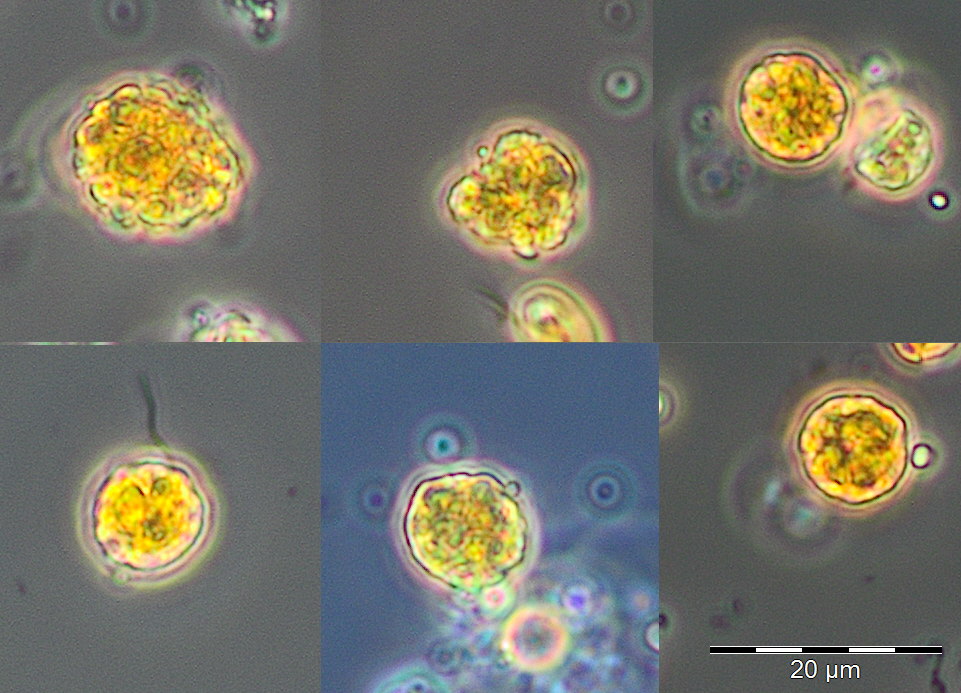
Am 8.5.2019 (Woche 19) begann die Periode des Sommer-Minimums der Phytoplankton-Biomasse. Die Kieselalge Rhizosolenia setigera dominierte (am 8.5.2019 mit 71 mg/m3). Daneben waren Plagioselmis prolonga, Hemiselmis sp. und Heterocapsa rotundata erwähnenswert, aber Mesodinium rubrum war fast vollständig verschwunden. Ein Übersichtsbild vom 22.5.2018 mit Cryptophyceen, Prymnesiales und Einzellern < 2µm (Picoplankton) wird in Bild 3 gezeigt. Am 30.5.2019 (Woche 22) trat kurzzeitig Dictyocha speculum mit 83 mg/m3 auf, das in manch anderen Jahren (2008, 2011, 2014, 2015, 2017) im späten Frühjahr große Biomassen bilden konnte (Bild 4).
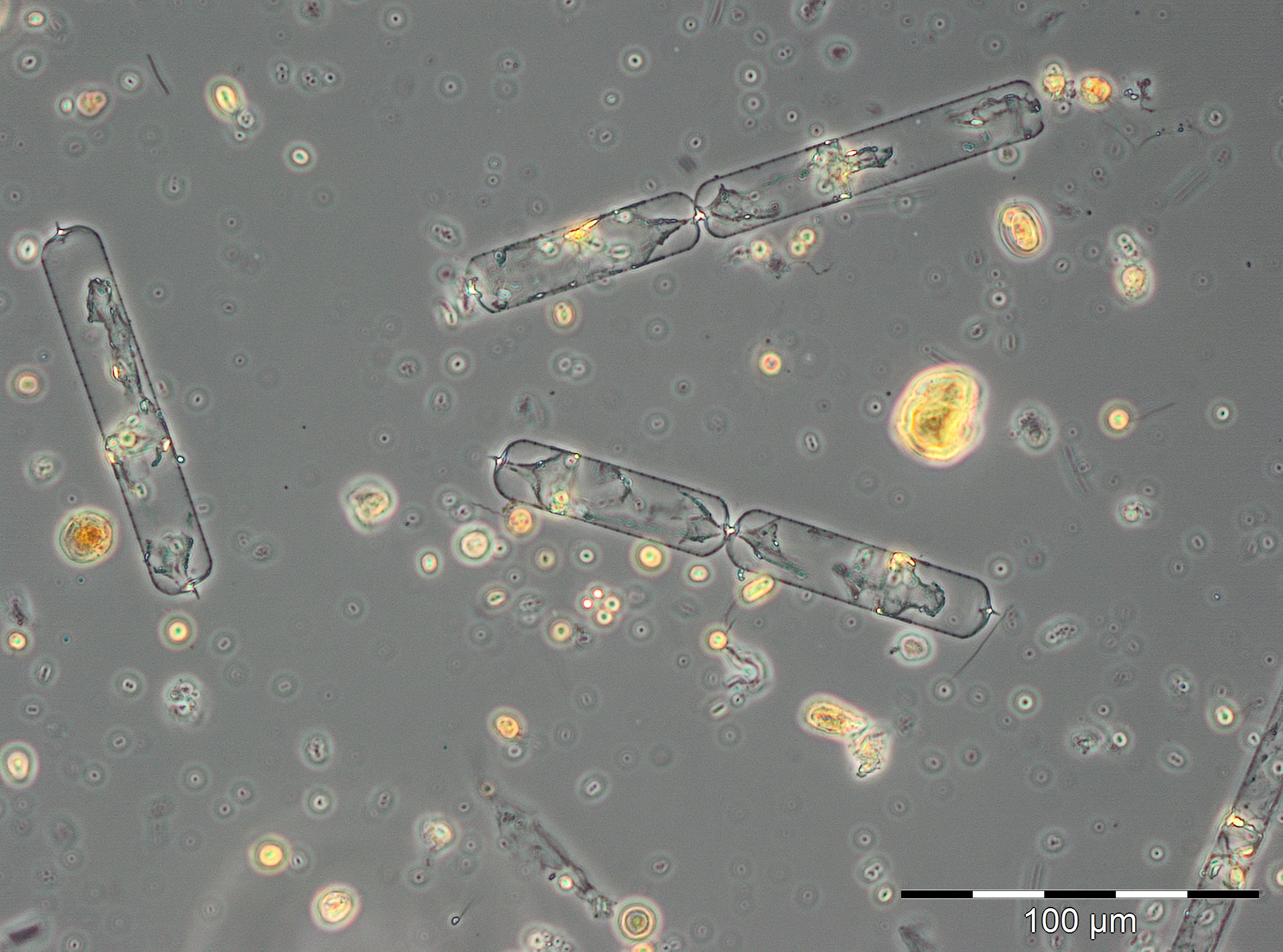
Die Entwicklung einer Sommerblüte von Kieselalgen begann am 19.6.2018 (Woche 25), unüblicherweise zu Beginn durch die Frühjahrsart Skeletonema marinoi dominiert (120 mg/m3). Am 26.6.2018 war Skeletonema marinoi aber fast wieder verschwunden, und es trat die typische Sommer-Kieselalge Dactyliosolen fragilissimus mit 98 mg/m3 auf. Diese wuchs zum 3.7.2018 auf 1916 mg/m3 an (Bild 5). Derart große Kieselalgen haben relativ wenig Zellplasma (wie Bild 5 zeigt) und entsprechend geringen Chlorophyll-Gehalt (vgl. Abb. 1). Als Begleitarten sind erwähnenswert: Ceratium tripos, Nodularia spumigena, Cymbomonas tetramitiformis, Alexandrium pseudogonyaulax und unbestimmte Prymnesiales. Die drastisch reduzierte Biomasse in Woche 28 ist durch schlechte Zählbarkeit der Probe wegen starker Sedimentbeimischungen oder durch Ankunft eines Wasserkörpers mit abweichendem Entwicklungsstand der Blüte an der Mess-Station hervorgerufen. Auf jeden Fall setzte sich die Kieselalgenblüte vom 17.7. bis 31.7.2018 (Woche 29-31) fort mit Biomassen von Dactyliosolen fragilissimus um 6000 mg/m3.
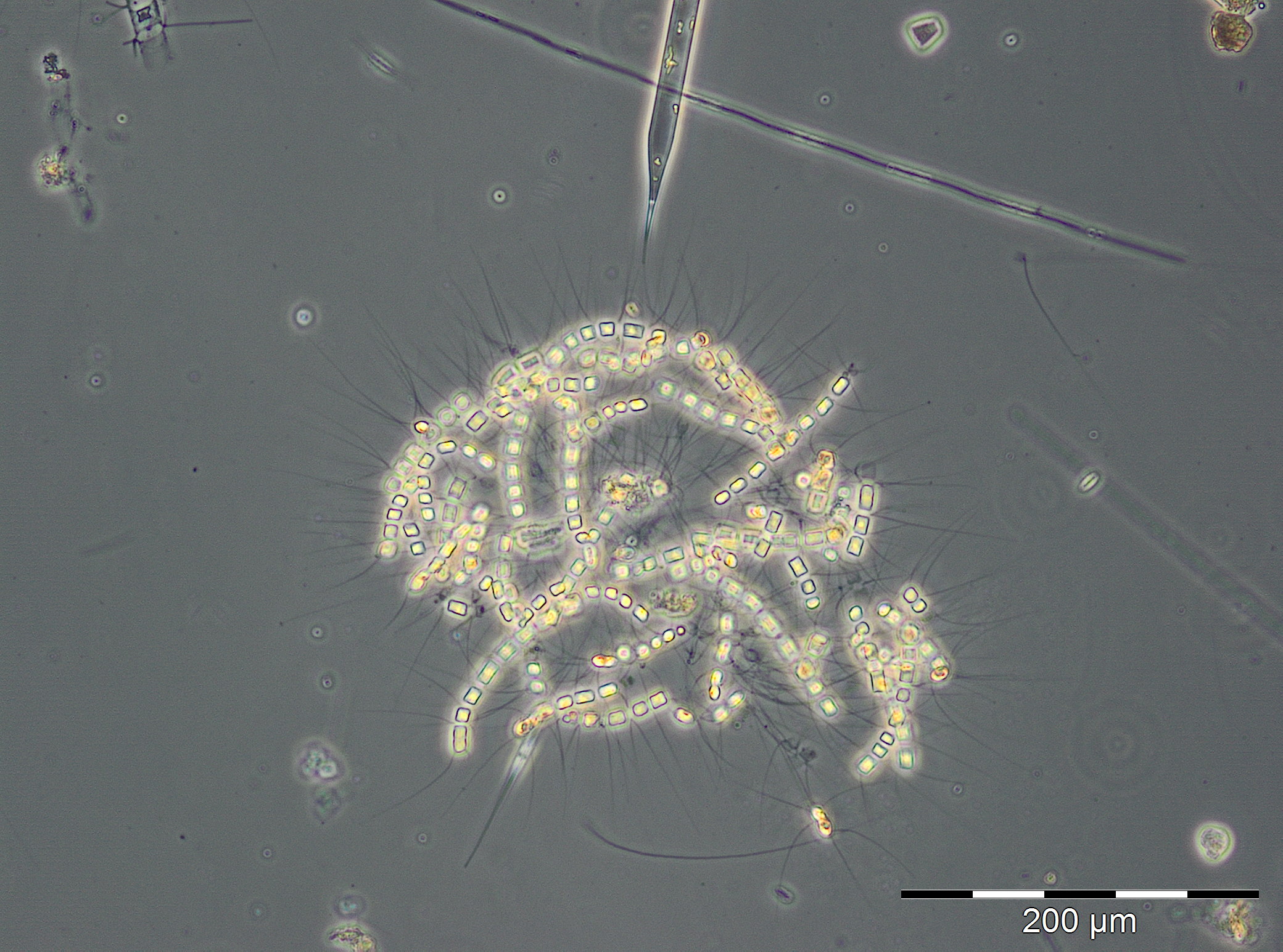
Interessant ist das kurzzeitige Auftreten hoher Biomassen von Nodularia spumigena (567 mg/m3) am 17.7.2018. Biomassen über 200 mg/m3 gelten nach Wasmund (1997) bereits als Blüte. Der Befund darf aber nicht überbewertet werden, da die aufschwimmenden Cyanobakterien-Matten gerade in Küstenbereichen kurzzeitig durch Wind und Strömung zusammengetrieben werden können, sich aber auch schnell wieder verteilen. An der polnischen Küste mussten wegen der giftigen Algenteppiche mehrere Strandabschnitte für Badende geschlossen werden („Zeit online“ vom 25.7.2018). An den deutschen Außenküsten bestand keine Gefahr. Am 17.7.2018 waren auch Dinoflagellaten bedeutsam: Prorocentrum cordatum (216 mg/m3), Ceratium tripos (195 mg/m3) und Alexandrium pseudogonyaulax (178 mg/m3).

Nach dem Zusammenbruch der Kieselalgen-Sommerblüte trat am 14.8.2018 (Woche 33) spontan wieder Mesodinium rubrum mit 405 mg/m3 auf. Die Proben von Wochen 34 und 35 waren schwer zählbar; sie enthielten viel Sediment und sehr viel Picoplankton. Zum 4.9.2018 (Woche 36) hatten sich wieder Kieselalgen entwickelt, jetzt aber nicht Dactyliosolen fragilissimus, sondern Pseudosolenia calcar-avis (316 mg/m3; Bild 6), Chaetoceros-Arten (C. contortus, C. circinalis, C. affinis, C. curvisetus, C. laciniosus, C. socialis; Bilder 6-8), Ditylum brightwellii, Cerataulina pelagica und Thalassiosira spp., aber auch Dinophyceen (Polykrikos schwartzii, Dinophysis acuminata, Ceratium spp.), Euglenophyceen (Eutreptiella) und Mesodinium rubrum.
Die Artenzusammensetzung änderte sich wieder zum 18.9.2018 (Woche 38). Neben dem erneuten Auftreten der Sommer-Kieselalge Dactyliosolen fragilissimus (124 mg/m3) kam nun auch die typische Herbst-Art Coscinodiscus granii hinzu (216 mg/m3). Chaetoceros spp., Pseudosolenia calcar-avis, Euglenophyceen, Mesodinium rubrum und die oben erwähnten Dinoflagellaten waren fast vollständig verschwunden. Die Probe vom 2.10.2018 (Woche 40) war stark versandet und die Biomasse wegen schwieriger Erkennbarkeit unterschätzt. Es zeigte sich aber eine Dominanz von Coscinodiscus granii. Diese Art trat aber am 9.10.2018 (Woche 41) gar nicht mehr auf. Es dominierte Pseudosolenia calcar-avis (219 mg/m3). P. calcar-avis etablierte sich erst seit 2010. Dessen starke Entwicklung wurde von Kaiser et al. (2016) analysiert. Lennoxia faveolata war in geringer Biomasse am 9.10.2018 präsent (Bild 9); es tritt erst seit 2009 sporadisch in unseren Proben auf (vgl. Wasmund et al. 2018).
Die Herbstblüte am 16.10.2018 (Woche 42) war wiederum durch eine andere Art bestimmt. Pseudosolenia calcar-avis war stark reduziert (78 mg/m3), aber Cerataulina pelagica hatte sich stark entwickelt (1909 mg/m3). Erwähnenswert ist auch das Auftreten des üblicherweise im Frühjahr vorkommenden Art Dictyocha speculum (57 mg/m3). Die nächste auswertbare Probe (8.11.2018, Woche 45) war von der Artenzusammensetzung wieder ganz anders, mit der Rangfolge, Thalassiosira spp., Ditylum brightwellii, Pseudo-nitzschia spp., Proboscia alata, Pseudosolenia calcar-avis, Cerataulina pelagica. Es war offensichtlich ein anderer Wasserkörper an die Station transportiert. Die Probe vom 13.11.2018 (Woche 46) passte wiederum zu der Probe von Woche 42: Cerataulina pelagica war absolut dominant (2129 mg/m3).

Die folgenden Proben waren nicht auswertbar (Bild 10). Auch der in Abb. 1 dargestellte Wert von Woche 49 dürfte wegen Versandung der Probe verfälscht sein. Erst die Probe vom 18.12.2018 (Woche 51) war unverfälscht. Auch hier dominierte immer noch Cerataulina pelagica (659 mg/m3), aber es waren andere bedeutsame Arten hinzugekommen: Pseudo-nitzschia spp. (202 mg/m3), Thalassiosira spp. (169 mg/m3), Ceratium tripos (140 mg/m3), Skeletonema marinoi (85 mg/m3), Mesodinium rubrum (44 mg/m3), Dactyliosolen fragilissimus (33 mg/m3).
Zusammenfassend ist festzustellen, dass die Frühjahrsblüte 2018 nur kurz war (etwa vom 6.3.2018 bis 17.4.2018), wie erwartet mit einer Sukzession von Kieselalgen (Skeletonema marinoi und Thalassiosira spp.) zu motilen Formen (Dinoflagellaten, Mesodinium rubrum). Die in vielen vorangegangenen Jahren dominante nackte Formen von Dictyocha speculum war kaum vorhanden. Vom 3.7. bis 31.7.2018 war eine typische Kieselalgenblüte (Dactyliosolen fragilissimus) sowie am 17.7.2018 eine kurzzeitige Cyanobakterienblüte vorhanden. Es schlossen sich ab September mehrere kleine Blüten unterschiedlicher Zusammensetzung an, dominiert durch Pseudosolenia calcar-avis (4.9.2018, 9.10.18) und Cerataulina pelagica (16.10.-18.12.2018). Die sonst häufig die Herbstblüte bildende Kieselalge Coscinodiscus granii und die typische Dinoflagellatengattung Ceratium waren nur schwach entwickelt.
Danksagung
Unser Dank gilt den Kolleginnen und Kollegen der Sektion Meereschemie (Lars Kreuzer, Jenny Jeschek, Birgit Sadkowiak), die die gemeinsamen Probenahmen begleiteten und die Nährstoffanalysen durchführten sowie den Kolleginnen und Kollegen der EDV (Dr. Steffen Bock, Dr. Susanne Feistel, Solvey Hölzel), die die technischen Voraussetzungen für die Arbeit mit den Daten und das Laden dieses Berichts auf die IOW-Seite absicherten.
Literatur
Kaiser, J., Belt, S.T., Tomczak, M., Brown, T.A., Wasmund, N., Arz, H.W., 2016: C25 highly branched isoprenoid alkenes in the Baltic Sea produced by the marine planktonic diatom Pseudosolenia calcar-avis. – Organic Geochemistry 93: 51-58.
Olenina, I., Hajdu, S., Andersson, A.,Edler, L., Wasmund, N., Busch, S., Göbel, J., Gromisz, S., Huseby, S., Huttunen, M., Jaanus, A., Kokkonen, P., Ledaine, I., Niemkiewicz, E. (2006): Biovolumes and size-classes of phytoplankton in the Baltic Sea. Baltic Sea Environment Proceedings No.106, 144pp.
http://www.helcom.fi/Lists/Publications/BSEP106.pdf
Wasmund, N. (1997): Occurrence of cyanobacterial blooms in the Baltic Sea in relation to environmental conditions. Int. Revue ges. Hydrobiol. 82: 169-184.
Wasmund N., Dutz J., Pollehne F., Siegel H. & Zettler M.L. 2018. Biological assessment of the Baltic Sea 2017. Meereswiss. Ber., Warnemünde 108: 1-101.
https://www.io-warnemuende.de/files/forschung/meereswissenschaftliche-berichte/mebe108_2018_assessment-bio.pdf
Wasmund, N., Topp, I., Schories, D. (2006): Optimising the storage and extraction of chlorophyll samples. Oceanologia 48: 125-144.
Autoren
IOW, 07.06.2019
Dr. Norbert Wasmund,
Susanne Busch,
Christian Burmeister.
Regina Hansen.
Leibniz Institute für Ostseeforschung Warnemünde (IOW),
Seestr. 15,
D-18119 Rostock-Warnemünde
Korrespondierender Autor: Dr. Norbert Wasmund

Der Zustand der Ostsee
- Jährliche Zustandseinschätzungen
- Fahrtberichte
- Daten der autonomen Messstationen
- Faktenblätter
- Kontinuierliche CO2- und O2-Messungen auf Frachtschiff FINNMAID
- Entwicklung der suboxischen und anoxischen Bereiche seit 1969
- Baltic Thalweg Transekte seit 2014
- Algenblüten vor Heiligendamm seit 1998
- Salzwassereinbruch Dezember 2014
- Salzwassereinbruch Januar 2003
- Barotrope Salzwassereinbrüche 1887 - 2018
- Weiterführende Literatur

IOW-Fotogalerie einzelliger Ostseealgen
... licht- und elektronenmikroskopische Aufnahmen pelagischer Mikroalgen