

Phytoplanktonentwicklung an der Küstenstation Heiligendamm im Jahre 2017
Die Sektionen Biologische Meereskunde und Meereschemie des Leibniz-Instituts für Ostseeforschung Warnemünde (IOW) führen wöchentliche Probennahmen an der Seebrücke Heiligendamm (54°08,76' N; 11°50,58' E) durch. In diesem Bericht werden die Ergebnisse der Phytoplankton-Untersuchungen vorgestellt. Es wurde Oberflächenwasser mit einer Pütz entnommen.
Die Phytoplanktonbiomasse wurde durch mikroskopische Zählung (UTERMÖHL-Methode) und die Chlorophyll-a-Konzentration durch Ethanol-Extraktion und fluorometrische Messung bestimmt. Dabei wurden die Methodenvorschriften von HELCOM zugrunde gelegt: http://www.helcom.fi/Documents/Action%20areas/Monitoring%20and%20assessment/Manuals%20and%20Guidelines/Guidelines%20for%20monitoring%20phytoplankton%20species%20composition,%20abundance%20and%20biomass.pdf Die Zählung der Phytoplankton-Proben erfolgte mit dem Zählprogramm OrgaCount und basiert auf der HELCOM-Artenliste, die jährlich aktualisiert wird: http://www.ices.dk/marine-data/vocabularies/Documents/PEG_BVOL.zip (Grundlagen siehe in Olenina et al. 2006). Die spezifischen analytischen Bedingungen der Chlorophyll-a-Bestimmung sind in Wasmund et al. (2006) dargelegt. Es werden hier entsprechend der Entscheidung der BLMP-Unter-Arbeitsgruppe Qualitätssicherung vom 11.9.2008 die Chlorophyll-a-Konzentrationen angegeben, die nicht für Phaeopigment korrigiert sind.

Die Phytoplanktonproben von 13 Messtagen konnten nicht mikroskopisch ausgewertet werden, da es sturmbedingt zu hohen Sedimentanteilen in den Proben gekommen war. Die Chlorophyllproben waren jedoch nicht betroffen, so dass sie Informationen über die Phytoplanktonbiomasse selbst bei fehlenden Phytoplanktonanalysen zuließen.
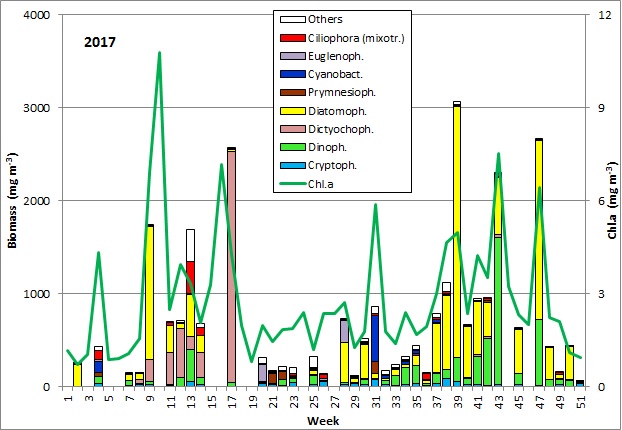
Die Ergebnisse sind in Abb. 1 dargestellt.
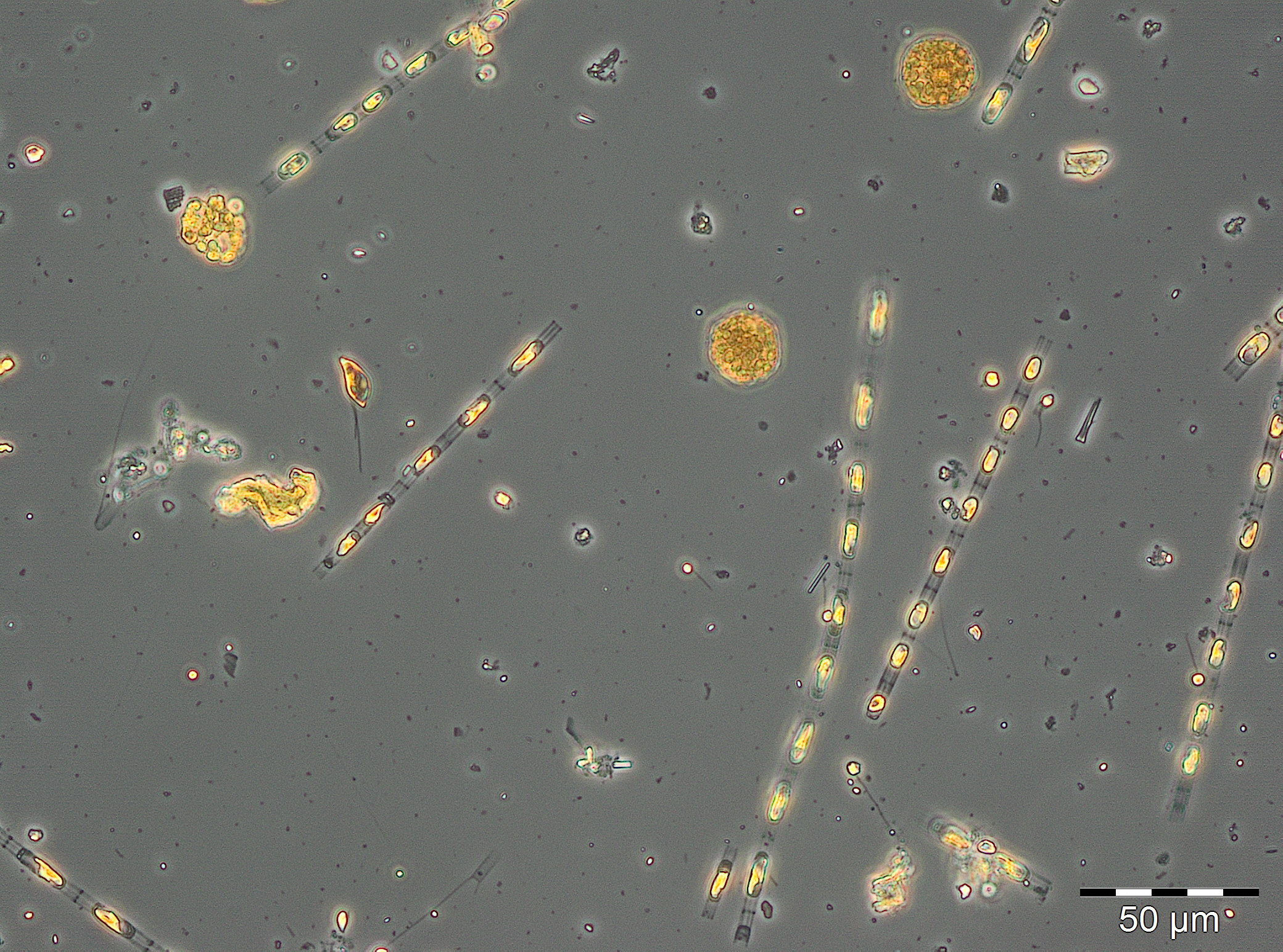
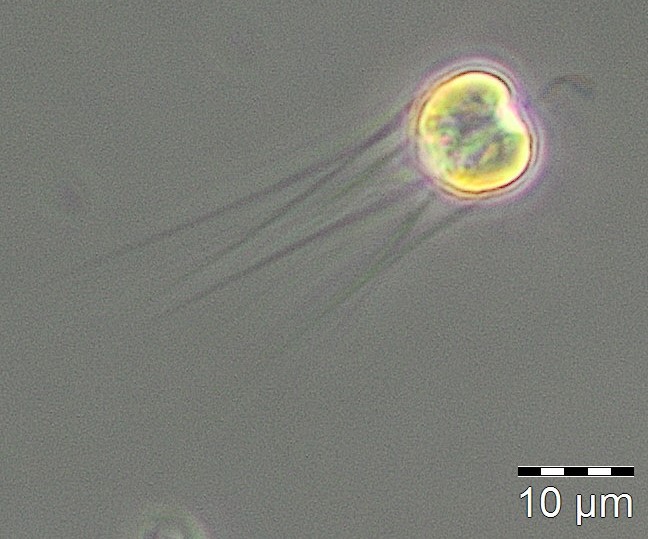
Der Jahresbeginn war gekennzeichnet durch die Kieselalge Rhizosolenia setigera, die bereits am 13.12.2016 mit einer Biomasse von 230 mg/m3 auftrat und bis zum 10.1.2017 (Woche 2) eine ungewöhnlich hohe Biomasse von 133 mg/m3 behielt. Noch ungewöhnlicher waren der hohe Chlorophyllwert und das sehr diverse Phytoplankton am 24.1.2017 (Woche 4). Außergewöhnlich für diese Jahreszeit war die hohe Biomasse (125 mg/m3) des Cyanobakteriums Aphanizomenon sp. (Bild 1), das normalerweise erst im Sommer und vorzugsweise in den nördlichen Teilen der Ostsee Blüten bildet. Es trat nun fast ausschließlich ohne Heterocyten auf, hatte aber meistens die hyalinen Endzellen, die ein Bestimmungsmerkmal sind. Probleme bei der Artansprache der Ostsee-Art sind bekannt (vgl. Laamanen 2002), deshalb allgemein nur als „Aphanizomenon sp.“ bestimmt. Das Fehlen von Heterocyten bei Anwesenheit von Nitrat (3,6 µM am 24.1.2017) wurde öfter berichtet (Laamanen 1996, Adams & Duggan 1999, Palińska & Surosz 2008). Das Absinken des Salzgehaltes von 17,5 psu am 10.1.2017 auf 10,3 psu am 24.1.2017 könnte ein Indiz für kurzzeitig eingeleitetes Süßwasser aus dem Conventer See sein, welches die hohen Aphanizomenon-Biomassen enthielt. In dem Falle wäre es nicht die Brackwasserart der Ostsee, sondern die typische Süßwasserart Aphanizomenon flos-aquae. Was gegen die Hypothese der Einleitung von Süßwasser spricht, wäre das Fehlen weiterer typischer Süßwasserarten. Diese könnten jedoch bei der Einmischung in das Brackwasser abgestorben sein, während Aphanizomenon sp. sehr salztolerant ist. Die wesentlichen Begleit-Taxa waren Mesodinium rubrum (41 mg/m3), Heterocapsa rotundata (41 mg/m3), Prymnesiales (38 mg/m3), Teleaulax spp. (21 mg/m3) und Eutreptiella spp. (21 mg/m3). In der quantitativ nicht auswertbaren Probe vom 31.1.2017 trat Aphanizomenon sp. schon nicht mehr auf und der Chlorophyll-Gehalt war deutlich abgesunken.
Ausgehend von einer relativ geringen Biomasse am 21.2.2017 (Woche 8) kam es innerhalb einer Woche zu einem gewaltigen Wachstum. Die bereits am 21.2.2017 dominierenden Arten Skeletonema marinoi (52 mg/m3) und Dictyocha speculum (42 mg/m3) stiegen zum 28.2.2017 (Woche 9) auf eine Biomasse von jeweils 1303 mg/m3 und 238 mg/m3 an, was fast schon für ein exponentielles Wachstum spricht. Wie schon in den vorjährigen Berichten möchten wir aber darauf hinweisen, dass der starke Biomasseanstieg nicht wirkliches Wachstum innerhalb eines Wasserkörper sein könnte, falls ein anderer Wasserkörper, der schon eine höhere Biomasse enthielt, die Station erreicht hat.
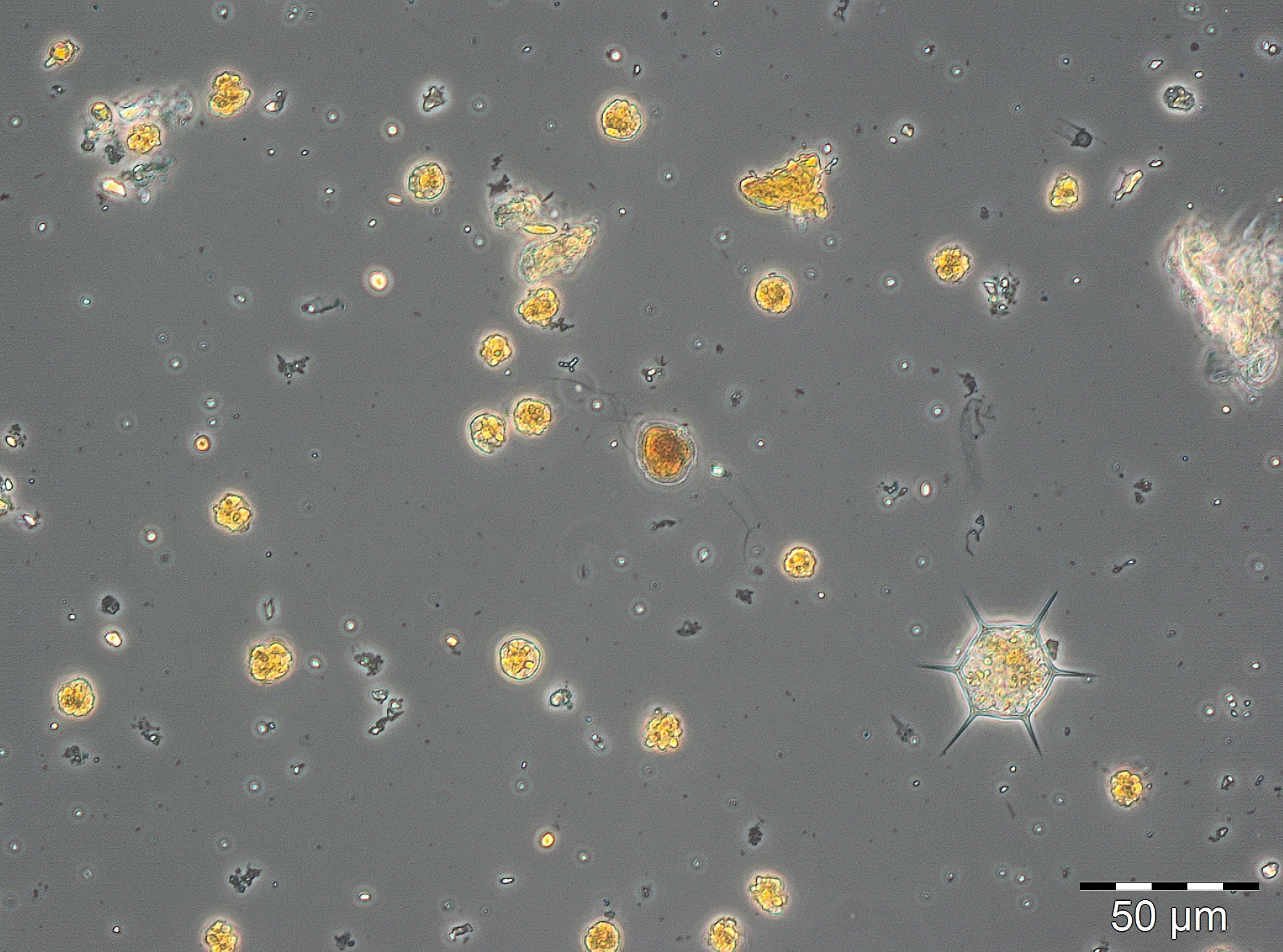
Der Höhepunkt der Frühjahrsblüte dürfte erst am 7.3.2017 (Woche 10) erreicht worden sein, was aus dem Spitzenwert des Chlorophyll a von 10,8 mg/m3 abzuleiten ist. Leider war die Phytoplanktonprobe vom 7.3.2017 mikroskopisch nicht auswertbar. In der Endphase dieser Blüte ging Skeletonema marinoi weiter zurück während Dictyocha speculum anstieg.
Am 28.3.2017 (Woche 13) zeigte sich eine deutlich veränderte Artzusammensetzung. Neben Skeletonema marinoi (407 mg/m3) und Dictyocha speculum (136 mg/m3) traten kurzzeitig hohe Biomassen des Ciliaten Mesodinium rubrum (343 mg/m3), des Dinoflagellaten Peridiniella danica (233 mg/m3) und der Chrysophycee Apedinella radians (313 mg/m³) auf. Bis zum vorjährigen Bericht wurde Dictyocha speculum in der grafischen Darstellung zu den Chrysophyceen gezählt, obwohl die Art neben anderen schon vor vielen Jahren aus den Chrysophyceen als eigenständige Klasse abgetrennt worden ist. Die eigentlichen Chrysophyceen waren bisher selten; deshalb wollten wir ab 2017 die Dictyochophyceen separat darstellen und die Chrysophyceen nicht gesondert in der Grafik ausweisen, sondern in die Restgruppe der „Sonstigen“ („Others“) stellen. Kurioserweise trat gerade in 2017 (aber nur am 28.3.2017) die Chrysophyceen mit Apedinella radians stark auf (Bild 3).

Anschließend wurde Dictyocha speculum wieder dominant und erreichte am 25.4.2017 (Woche 17) ein extremes Maximum von 2487 mg/m3 (Bild 4). Die Chlorophylldaten zeigten, dass das Maximum dieser zweiten Frühjahrsblüte bereits am 18.4.2017 (Woche 16) gefunden wurde, allerdings ohne auswertbare Phytoplanktonprobe. An dem Tag hat die Phosphatkonzentration ein Minimum von 0,03 µmol/l erreicht und stieg anschließend wieder an. Die Konzentrationen an Nitrat und Nitrit sanken zum 16.5.2017 (Woche 20) auf 0,05 µmol/l. An dem Tag dominierte Eutreptiella sp. mit 179 mg/m3; daneben sind die Cryptophyceen Teleaulax sp. und Plagioselmis prolonga sowie der Dinoflagellat Heterocapsa rotundata erwähnenswert. Damit ist die Frühjahrsphase beendet. Eine Spätfrühjahrsblüte von Dictyocha speculum nach der „klassischen“ Kieselalgen-Frühjahrsblüte wurde in manchen Jahren gefunden und ist durchaus typisch für dieses Seegebiet.
Während des Sommer-Minimums von Woche 21 bis 26 mit Nitrat+Nitrit-Konzentrationen von < 0,08 µM (außer einem Ausreißer) waren die Phytoplankton-Biomassen immer unter 250 mg/m3. Anfangs dominierten unbestimmte mixotrophe Prymnesiales (115 mg/m3), meistens in der Größenklasse von 2-4 µm. Sie wurden zunehmend von Gymnodiniales und später von Plagioselmis prolonga, Mesodinium rubrum und Heterocapsa rotundata verdrängt. Am 20.6.2017 (Woche 25) wurden die heterotrophen Nanoflagellaten Telonema sp. und Katablepharis remigera (in Abb. 1 als „Others“ erfasst) dominant.
Die Nitratkonzentrationen hatten sich zum 4.7.2017 (Woche 27) regeneriert (0.79 µM), vielleicht auch nur wegen windbedingter Aufwirbelung (Windstärke 6), denn die Planktonprobe war wegen hohen Sedimentanteils nicht auswertbar. Nach den starken Nordwestwinden mit Salzwassereinstrom (Salzgehalt am 4.7.2017: 12.7 psu)
traten am 11.7.2017 (Woche 28) die typische Sommer-Kieselalge Dactyliosolen fragilissimus (410 mg/m3) und die Euglenophycee Eutreptiella sp. (230 mg/m3) auf. Der folgende niedrige Biomassewert (Woche 29) wird durch unvollständiges Auszählen der Probe wegen starker Sedimentbeimischung bedingt sein. Jedenfalls ist am 25.7.2018 (Woche 30) Dactyliosolen fragilissimus mit 351 mg/m3 immer noch dominant.
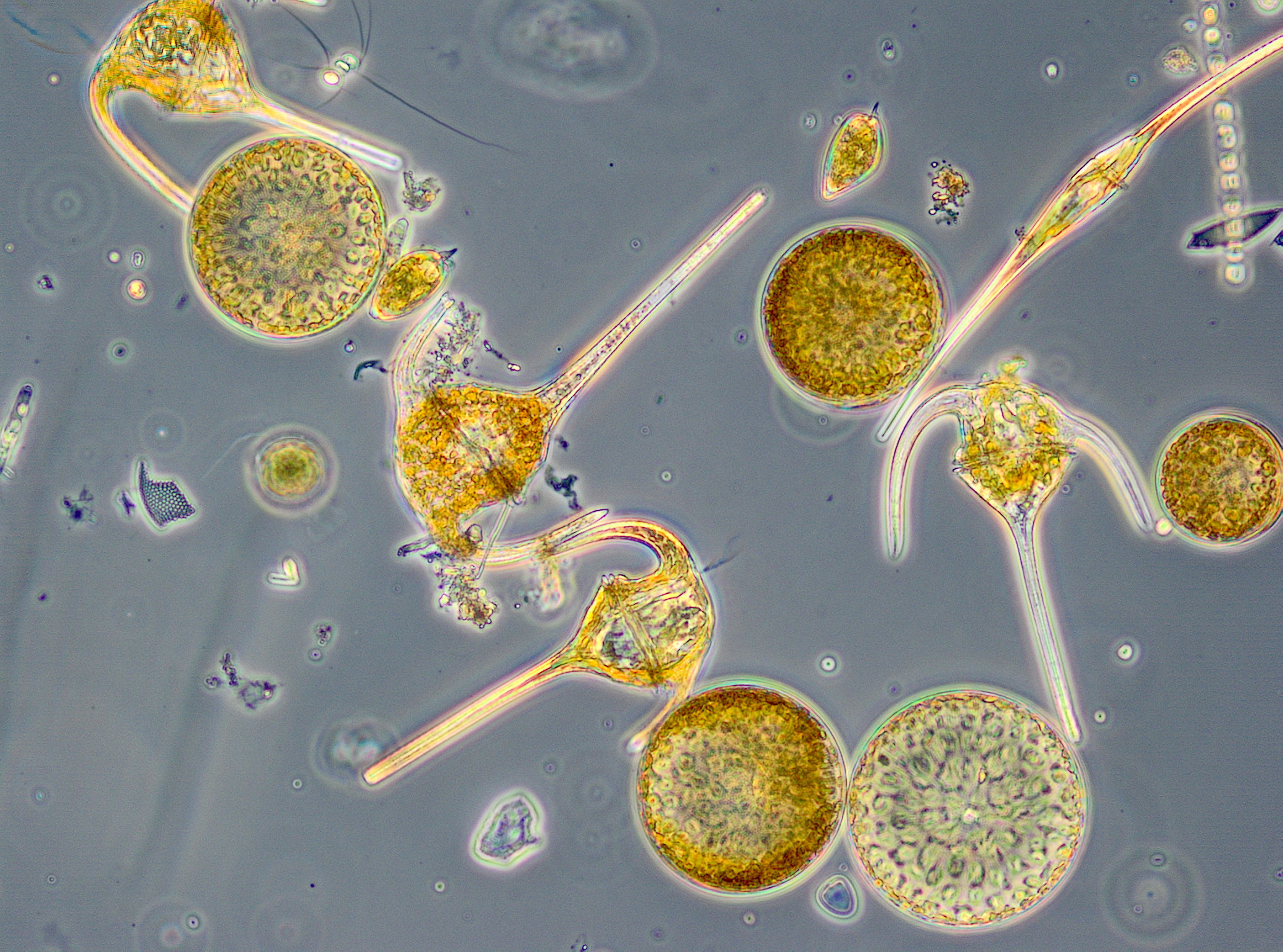
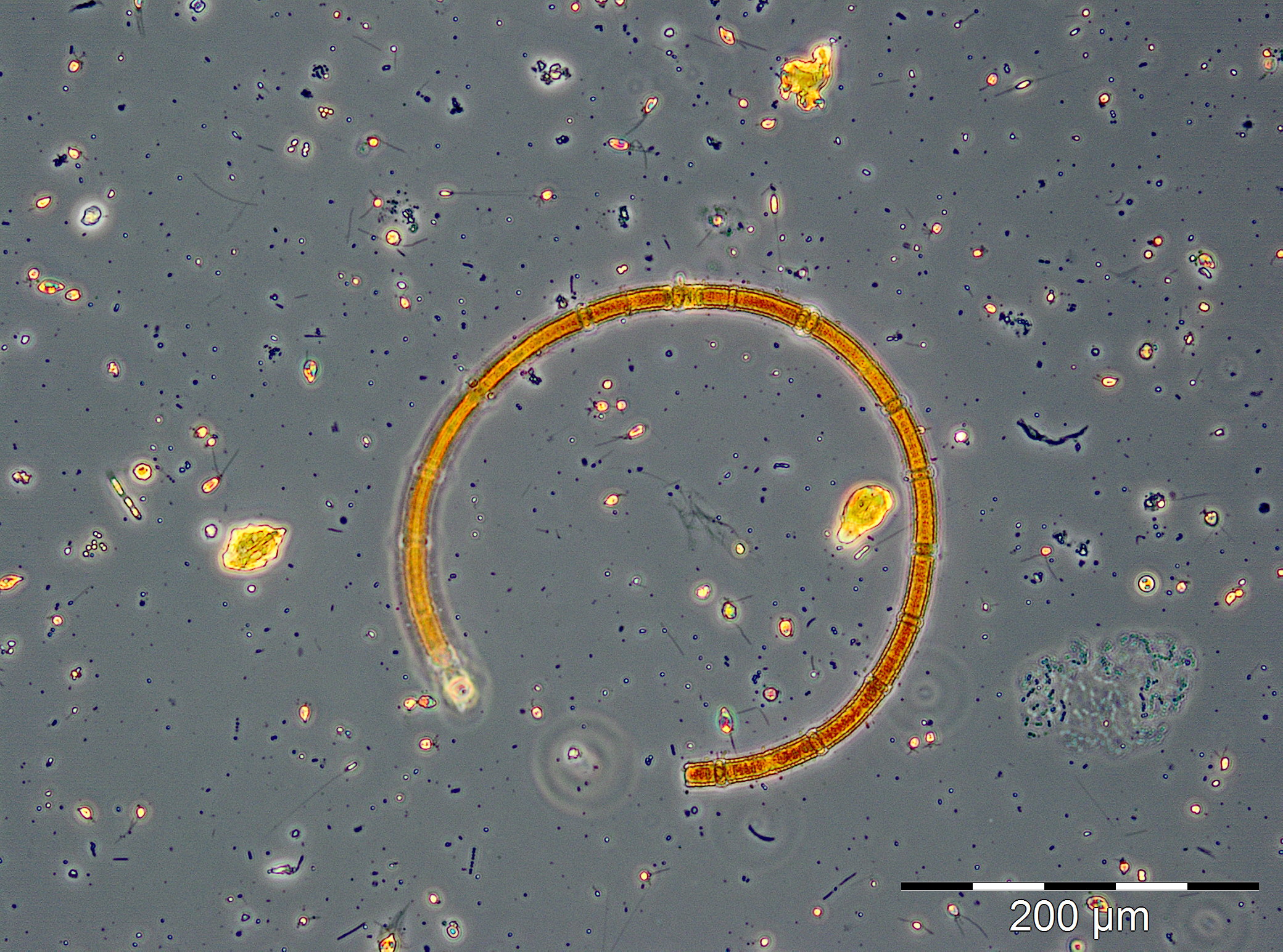
Zum 1.8.2017 (Woche 31) war Dactyliosolen fragilissimus fast ganz verschwunden und es erschienen spontan die stickstoff-fixierenden Cyanobakterien Aphanizomenon sp. mit 221 mg/m3 (Bild 5) und Nodularia spumigena mit 228 mg/m3 (Bild 6). Diese Cyanobakterienblüte war für dieses Seegebiet ungewöhnlich stark, aber nur von kurzer Dauer, wie schon im Jahre 2016. Bereits am 8.8.2017 (Woche 32) waren die Cyanobakterien fast vollständig verschwunden. An den folgenden 3 Messtagen (Woche 33-35) kamen langsam Ceratium tripos, Pseudosolenia calcar-avis, Alexandrium pseudogonyaulax, Prorocentrum cordatum und Prorocentrum micans zur Entwicklung. Ab dem 12.9.2017 (Woche 37) entwickelte sich nochmals eine Kieselalgenblüte. Sie war am 12.9.2017 geprägt von Dactyliosolen fragilissimus (374 mg/m3), am 19.9.2017 aber hauptsächlich von Cerataulina pelagica (419 mg/m3) und Coscinodiscus granii (308 mg/m3). Der Höhepunkt der Blüte wurde am 27.9.2017 (Woche 39) erreicht mit klarer Dominanz von Coscinodiscus granii (2451 mg/m3) und Ceratium tripos (213 mg/m3); Bild 7.
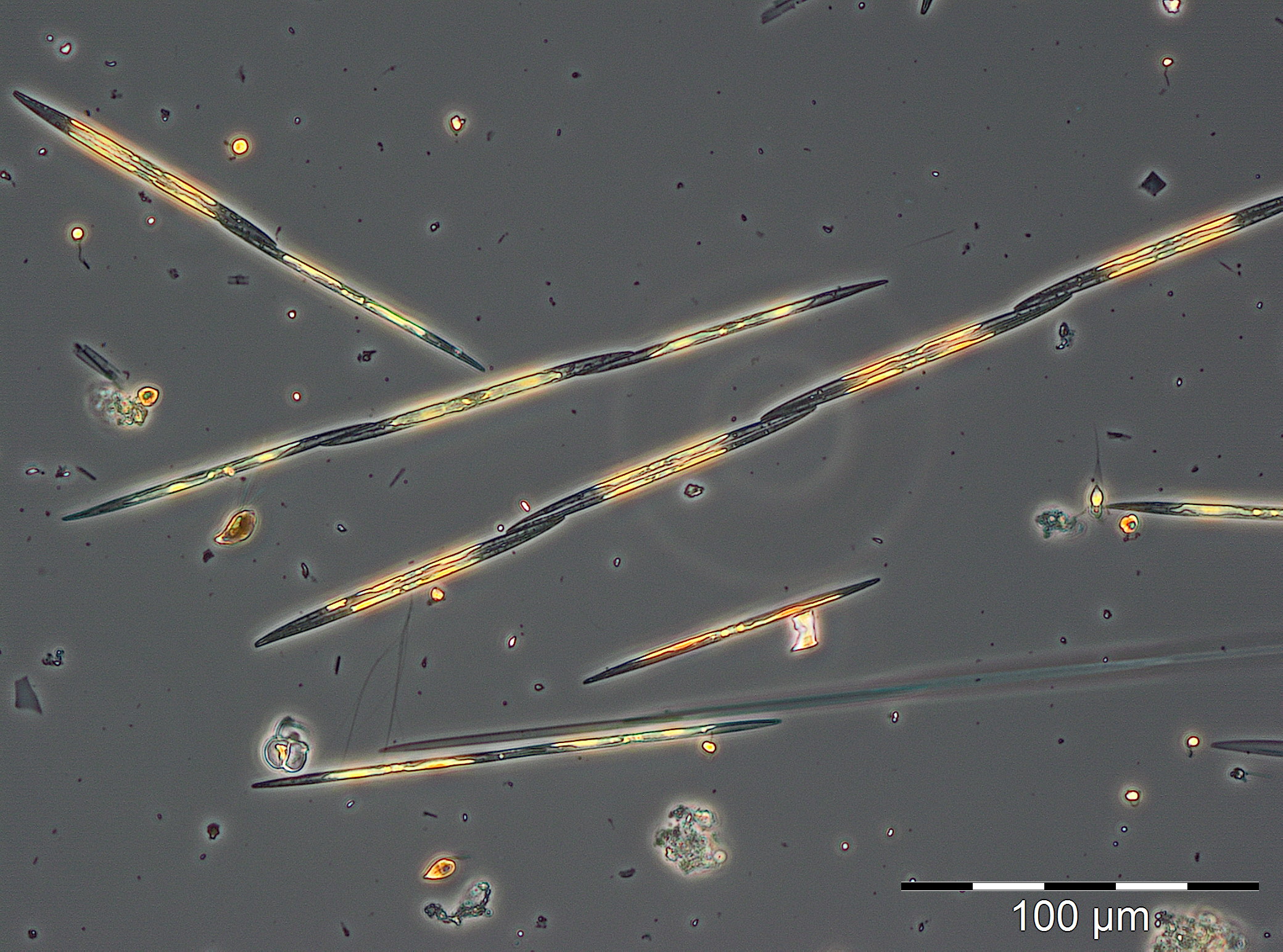
Coscinodiscus granii und Ceratium tripos sind typische Arten für die Herbstblüte, aber sie traten im vergangenen Jahr kaum auf. Es wurde bereits der Verdacht geäußert, dass Ceratium langfristig zurückgeht. Aber im Jahre 2017 entwickelte sich Ceratium tripos wieder stark. Es erreichte am 24.10.2017 (Woche 43) seinen Höhepunkt mit 1129 mg/m3. Dagegen war Coscinodiscus granii bis zum 24.10.2017 völlig verschwunden.
Stattdessen trat die potenziell toxische Kieselalgengattung Pseudo-nitzschia stark in Erscheinung. Sie erreichte am 21.11.2017 (Woche 47) ihren Höhepunkt mit 1221 mg/m3 (Bild 8). Elektronenmikroskopische Untersuchungen ergaben, dass es sich hauptsächlich um Pseudo-nitzschia pseudodelicatissima handelt, während Pseudo-nitzschia pungens weniger vertreten war (Bild 9). Eine Blüte dieser Gattung ist in der westlichen Ostsee selten. Pseudo-nitzschia kam bis zum 12.12.2017 (Woche 50) vor, während an dem Tag aber die Kieselalgen Rhizosolenia setigera und Thalassiosira spp. mit Biomassen von jeweils über 70 mg/m3 schon stärker entwickelt waren.

Zusammenfassend ist festzustellen, dass bereits im Januar ein ungewöhnliches Auftreten des Cyanobakteriums Aphanizomenon sp. beobachtet werden konnte. Die Frühjahrsblüte 2017 bestand aus zwei deutlich voneinander getrennten Phasen, was eine durchaus normale Sukzession darstellt. Die erste Phase war als fast reine Kieselalgenblüte (Skeletonema marinoi) ausgeprägt, während die zweite Phase üblicherweise aus Flagellaten besteht. Im Jahre 2017 war die Flagellatenphase durch die auch in vielen vorangegangenen Jahren dominante nackte Formen von Dictyocha speculum repräsentiert. Im Sommer war eine moderate Kieselalgenblüte (Dactyliosolen fragilissimus) sowie eine kurzzeitige, aber relativ starke Cyanobakterienblüte vorhanden. Geringe Cyanobakterien-Biomassen kamen noch bis Mitte September vor. Die Herbstblüte war lang anhaltend und wurde deutlich von Kieselalgen (Coscinodiscus granii, gefolgt von Pseudo-nitzschia pseudodelicatissima) dominiert. Auch die für den Herbst typische Dinoflagellatengattung Ceratium war im Gegensatz zu den Vorjahren wieder blütenbildend.
Danksagung
Unser Dank gilt den Kolleginnen und Kollegen der Sektion Meereschemie (Lars Kreuzer, Jenny Jeschek, Birgit Sadkowiak), die die gemeinsamen Probenahmen begleiteten und die Nährstoffanalysen durchführten sowie den Kolleginnen und Kollegen der EDV (Dr. Steffen Bock, Dr. Susanne Feistel, Solvey Hölzel), die die technischen Voraussetzungen für die Arbeit mit den Daten und das Laden dieses Berichts auf die IOW-Seite absicherten.
Literatur
Adams, D.G., Duggan, P.S. (1999): Tansley Review No. 107: Heterocyst and akinete differentiation in cyanobacteria. New Phytol. 144: 3-33.
Laamanen M. (1996): Cyanoprokaryotes in the Baltic Sea ice and winter plankton. Algological Studies 83: 423-433.
Laamanen M. (2002): Genetic and species diversity of planktonic cyanobacteria in the northern Baltic Sea. Finnish Institute of Marine Research, Contributions No. 4.
Olenina, I., Hajdu, S., Andersson, A.,Edler, L., Wasmund, N., Busch, S., Göbel, J., Gromisz, S., Huseby, S., Huttunen, M., Jaanus, A., Kokkonen, P., Ledaine, I., Niemkiewicz, E. (2006): Biovolumes and size-classes of phytoplankton in the Baltic Sea. Baltic Sea Environment Proceedings No.106, 144pp.
http://www.helcom.fi/Lists/Publications/BSEP106.pdf
Palińska, K.A., Surosz, W. (2008): Population of Aphanizomenon from the Gulf of Gdańsk (Southern Baltic Sea): differences in phenotypic and genotypic characteristics. Hydrobiologia 607: 163-173.
Wasmund, N., Topp, I., Schories, D. (2006): Optimising the storage and extraction of chlorophyll samples. Oceanologia 48: 125-144.
Wasmund, N., Siegel, H., Bohata, K., Flohr, A., Hansen, A., Mohrholz, V. (2015): Phytoplankton stimulation in frontal regions of Benguela upwelling filaments by internal factors. Frontiers in Marine Science 3 (210): 1-17. doi: 10.3389/fmars.2016.00210.
Autoren
IOW, 08.06.2018
Dr. Norbert Wasmund,
Susanne Busch,
Christian Burmeister.
Regina Hansen.
Leibniz Institute für Ostseeforschung Warnemünde (IOW),
Seestr. 15,
D-18119 Rostock-Warnemünde
Korrespondierender Autor: Dr. Norbert Wasmund

Der Zustand der Ostsee
- Jährliche Zustandseinschätzungen
- Fahrtberichte
- Daten der autonomen Messstationen
- Faktenblätter
- Kontinuierliche CO2- und O2-Messungen auf Frachtschiff FINNMAID
- Entwicklung der suboxischen und anoxischen Bereiche seit 1969
- Baltic Thalweg Transekte seit 2014
- Algenblüten vor Heiligendamm seit 1998
- Salzwassereinbruch Dezember 2014
- Salzwassereinbruch Januar 2003
- Barotrope Salzwassereinbrüche 1887 - 2018
- Weiterführende Literatur

IOW-Fotogalerie einzelliger Ostseealgen
... licht- und elektronenmikroskopische Aufnahmen pelagischer Mikroalgen